
| Hours post-fertilization | Control (FSW) | Low (0.01 mg/L) | Mid (0.1 mg/L) | High (1 mg/L) |
|---|---|---|---|---|
| 4 | 27.0 ± 3.9 | 27.3 ± 4.9 | 24.0 ± 5.4 | 26.0 ± 3.2 |
| 9 | 12.4 ± 7.5 | 12.6 ± 8.8 | 12.9 ± 6.1 | 11.1 ± 8.0 |
| 14 | 11.3 ± 5.6 | 12.7 ± 4.7 | 14.2 ± 7.2 | 9.6 ± 4.6 |
1 Introduction
Petroleum-derived plastic pollution is a rapidly escalating environmental concern (Stubbins et al. 2021). Global plastic production has doubled over the last decade, reaching an estimated 400 million tons in 2026. Of this, approximately 8-13 million tons of mismanaged plastic waste end up in the oceans annually, adding to an estimated 200 million tons already present in marine environments (Jambeck et al. 2015). Plastic pollution is increasingly concentrated in marine biodiversity hotspots, where major inputs from coastal regions in the Indo-Pacific overlap with coral reef ecosystems of exceptional ecological value, including the Coral Triangle. Ocean currents also transport and accumulate this debris in large offshore systems such as the Great Pacific Garbage Patch, highlighting how regional and coastal plastic pollution can be both a local and a global issue.
Once in the ocean, plastics are exposed to UV radiation and mechanical weathering causing them to become brittle and fragment into microplastics (plastics < 5 mm) and nanoplastics (plastics <1 mm) (MacLeod et al. 2021). As plastics break down physically, they also undergo chemical degradation, releasing chemical additives and breakdown products directly into coastal waters as leachate (Hahladakis et al. 2018). These leachates originate from multiple sources, including landfills, sewage, and nonpoint-source runoff, and ultimately concentrate in urban coastal zones. Whether in marine or terrestrial environments, plastic debris can release more than 10,000 chemical compounds. There is growing public awareness and concern over the impacts of plastic-derived leachate on human and environmental health.
Plastic leachate is a complex chemical mixture containing additives and degradation byproducts, such as UV stabilizers, colorants, plasticizers, bisphenol A (BPA), and per- and polyfluoroalkyl substances (PFAS) (Rochman et al. 2019). Among these, plasticizers (especially phthalates or phthalate acid esters (PAEs) used to enhance plastic flexibility), are the most abundant, accounting for up to 70% of the mass of PVC products. Phthalates are endocrine-disrupting chemicals (EDCs) that can mimic or interfere with hormonal signaling pathways that regulate developmental, reproductive, and immune function in both humans and wildlife (Maqbool et al. 2016). These additives are increasing in the marine environment alongside overall plastic pollution.
Early life stages in marine invertebrates (embryos and larvae) are often especially vulnerable to pollution (Richmond et al. 2018). Coral embryos are lipid-rich, and may be particularly susceptible to phthalates and other EDCs which are lipophilic. While endocrine signaling in cnidarians is not fully understood, endocrine hormones are known to play important roles in coral spawning timing, and embryonic cellular development (Tarrant et al. 2004; Tarrant 2005; Tarrant 2007). There is potential for plastic-derived leachate endocrine-like disruption, and if coral sexual reproduction and recruitment is negatively impacted, whole coral reef ecosystems could be degraded over time (Richmond 1997).
Relatively few studies have investigated the impacts of plastic-derived leachate on marine invertebrates. Studies that have been conducted primarily measure affects on fertilization, survival, development, larval motility, settlement, and metamorphosis. Phthalates and leachates from weathered polyvinyl chloride (PVC) affect larvae in the sea urchins Paracentrotus lividus (Gambardella et al. 2024; Jimenez-Guri et al. 2023; Rendell-Bhatti et al. 2021) and Strongylocentrotus purpuratus (Paganos et al. 2023). Embryonic development of the brown mussel, Perna perna, is sensitive to microplastic leachate (Gambardella et al. 2024). Even leachate from bioplastics contain additives that reduce egg fertilization, larval motility, and survival during the early development of the mussel Mytilus galloprovincialis (Capolupo et al. 2023). In the stony coral Stylophora pistillata, high exposure levels of dimethyl phthalate (DMP, 100 ug/L) reduced larval settlement, while exposure to environmentally relevant levels of plastic additive (1 ug/L of 4-nonylphenol 4-NP) reduced settlement of the soft coral Rhytisma fulvum (Vered and Shenkar 2022). (Berry et al. 2019) found limited effects of weathered polypropylene on fertilization of the broadcast spawning coral, Acropora tenuis, but no significant effects on embryo development and larval settlement. Wilkins et al. (2024) found no reduced fertilization rates in Montipora capitata exposed to microplastics and leachates from commercially sources virgin polypropylene (PP), high density polyethylene (HDPE), and low-density polyethylene (LDPE) microspheres. (Wilkins and Richmond 2025) found delayed or cumulative effects of reduced planulae survival and settlement in Montipora capitata and Porites corals exposed to virgin LDPE and HDPE leachates, while HDPE promoted settlement.
Together these studies indicate that leachate may be more toxic than physical particles, and weathered plastics are more toxic than virgin plastics. They also highlight the complexity of studying plastic-derived leachate and how our understanding of the current impacts of plastic leachate on marine organisms remains limited for several reasons. Plastic leachates are complex mixtures of thousands of additives whose composition varies with polymer type, age, and weathering state, making it difficult to standardize exposure treatments (Gunaalan et al. 2020). Many leachate compounds occur at very low environmental concentrations (ng-ug/L range), and their detection and quantification require highly sensitive analytical techniques, specialized expertise, and advanced infrastructure (Saliu et al. 2020). In addition, many past experimental studies still rely on high, acute exposure concentrations, which limit physiological interpretation in an environmentally relevant context (Weis and Palmquist 2021). Finally, integrative physiological studies remain scarce, with most research focusing on survival, growth, or gross developmental endpoints rather than mechanistic responses such as gene expression or microbiome dynamics.
In this study, we chose to focus on exploring the potential for endocrine-like disruption and non-linear responses in early life history stages of coral. We chose PVC plastic because it has a high phthalate content. We examined how PVC microplastic leachate impacts the development, microbiome, and transcriptomic signatures of embryos of the coral Montipora capitata. Coral reefs are one of the most valuable ecosystems on the planet; despite covering less than 1% of the oceans, they support at least 25% of all marine species (Reaka-Kudla 1997). Reef-building corals provide global services estimated to be worth US$2.7 trillion per year by protecting coasts from storm surge and wave erosion, supporting tourism and fisheries, and contributing to medicinal compound discovery (Souter et al. 2020). However, coral reefs are already in decline with global cover reduced by around 50% since the 1950s, and vulnerable to increasing coastal marine plastic pollution (Eddy et al. 2018, 2021; Oppen et al. 2017). Plastic pollution is ubiquitous across coral reef ecosystems globally (Pinheiro et al. 2023) as they are located near major coastal sources of plastic input and are influenced by the same oceanographic systems that transport and concentrate marine debris (Lamb et al. 2018). One study found that plastic debris correlated with higher rates of disease (Lamb et al. 2018), and another showed that poorer water quality, due to closer proximity to an urban center, reduced recovery after a heatwave-induced bleaching event (Claar et al. 2020).
M. capitata is a broadcast spawner that releases gametes into the water column during synchronous spawning events. Fertilization and embryonic development occur in the water column before larvae settle and undergo metamorphosis. Early life history stages can be particularly vulnerable to plastic leachates due to direct exposure to surrounding seawater and their high sensitivity during critical periods of cellular differentiation (Zhang et al. 2021; Lynch et al. 2022).
Given that PVC leachates contain endocrine disrupting chemicals with potential to induce low-dose effects at environmentally relevant concentrations, we hypothesize that PVC leachate exposure would produce low-dose effects across organismal, molecular, and holobiont responses. Specifically, we predicted that PVC leachate exposure would:
H1 Reduce embryo survival and impair normal development
H2 Alter gene expression in pathways required for early embryonic development
H3 Disrupt the microbiome community and microbial function
This study provides insight into how plastic-derived chemical pollution may affect coral early life stages, a critical but under-explored phase of reef resilience and recovery.
2 Methods
2.1 Gamete collection and ecotoxicity assay exposure
The rice coral, Montipora capitata, is an ecologically important reef builder, and is a hermaphroditic synchronous split-spawner that releases buoyant egg-sperm bundles during the summer months (Padilla-Gamiño and Gates 2012). In our study, we collected coral colonies on June 30th, 2024 from Kāneʻohe Bay, on the east shore of Oʻahu, Hawaiʻi. Kāneʻohe Bay is a 60 km2 embayment with several riverine inputs, numerous fringing and patch reefs, and a large barrier coral reef rimming it. Under the State of Hawai‘i Division of Aquatic Resources Special Activities Permit 2024-35, we collected 20 genetically distinct M. capitata coral colonies (each approximately 30 cm in diameter) via snorkel from Patch Reef #1 at 21.431619°N, 157.787127°W.
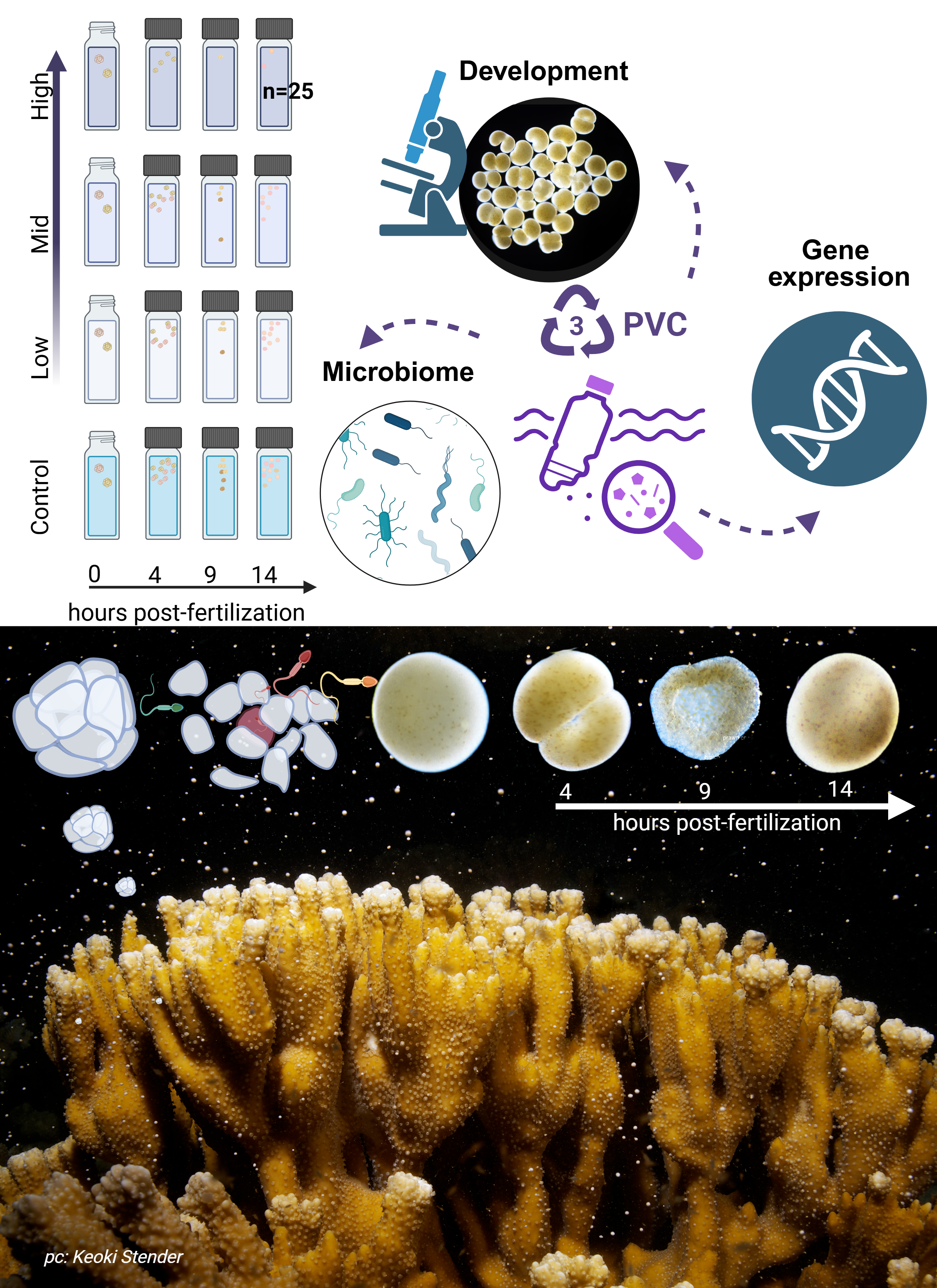
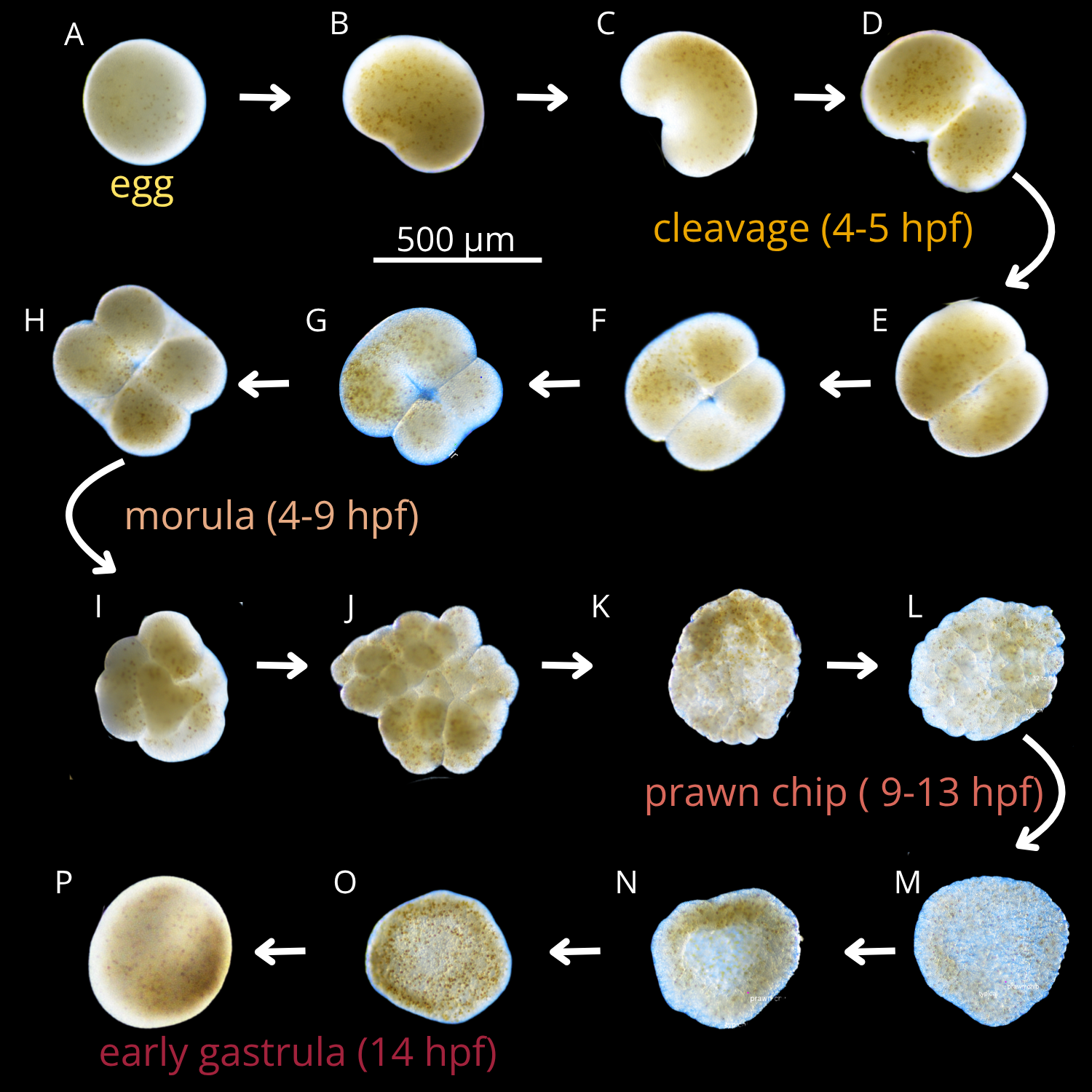
Immediately after collection, the coral colonies were carefully transported to the Hawaiʻi Institute of Marine Biology (HIMB), placed in two large 650 L holding tanks with flow-through seawater, and allowed to acclimate for 5 days prior to the spawning event during the new moon (Padilla-Gamiño et al. 2011). Egg-sperm bundles of Montipora capitata were collected across three consecutive nights (5th, 6th, and 7th of July 2024, on the new moon and 2 days after). On each night prior to spawning (~19:00 HST), coral colonies were isolated in individual 12-quart (11.36 L) containers surrounded by a flow-through seawater bath to maintain stable ambient seawater temperatures (26.3 ± 0.3 °C). Sixteen out of the twenty parent colonies spawned over the three-day event (Figure 16). We collected egg-sperm bundles from spawning colonies using transfer pipettes and deposited them into labeled 50 mL Falcon tubes containing 30 mL of 0.22 μm-filtered seawater (FSW). In the 1-hour window after egg-sperm bundle release and before bundle breakup, sperm release, and egg hydration, we performed controlled fertilizations by combining one intact bundle from each of two genetically distinct colonies in a bundle-bundle cross (Figure 1). Only intact bundles were used. Crosses were conducted in 20 mL scintillation vials containing 20 mL of either 0.22 μm filtered seawater (control) or one of the three PVC leachate treatments. Fertilized bundles were incubated on a laboratory bench in the dark at 26.8 ± 0.3°C for either 4, 9, or 14 hours post-fertilization (hpf). In Montipora capitata, these time points correspond to key developmental stages: 4 hpf (initial cleavage Figure 2 B-D), 9 hpf (prawnchip stage Figure 2 K-N), and 14 hpf (early gastrulation Figure 2 O,P). The experiment included 4 PVC leachate levels and 3 developmental stages, and resulted in 300 individual bundle-bundle crosses (Figure 1). A subset of 120 crosses (40 per stage) was fixed in 1.5 mL snap-cap micro-centrifuge tubes with 1 mL of 4% Z-fix (Anatech Ltd.) for assessment of survival, morphology, and developmental progression via microscopy. The remaining 180 crosses (60 per stage) were preserved in 500 μL of DNA/RNA ShieldTM (Zymo Research, USA) and stored at -80°C for subsequent DNA and RNA extraction to investigate microbial community composition and gene expression responses.
2.2 PVC leachate preparation, dilution, and analysis
PVC microplastics were obtained following the procedure described by Isa et al. (2024). Briefly, PVC granulates ≤ 500 μm were obtained by grinding PVC pellets, and polymer identity was confirmed by FTIR analysis (see supplementary material X). The material was then subjected to artificial weathering in a QUV accelerated weathering tester (Q-LAB, Saarbrucken, Germany) using UV lamps at 340 nm, with 0,76 W m−2 nm−1. The aging test was conducted at 65°C for a total time of 460 h. The total UV dose administered (measured by a bolometer) was 170 MJ/m2. This dose roughly corresponds to 10 months of outdoor exposure at 31°N latitude (Isa et al. 2025) to simulate photo-degradation processes experienced by plastic debris in the marine environment.
We prepared the PVC leachate by adding 250 mg of UV-weathered PVC microplastics to 250 mL of 0.22 μm-filtered seawater (FSW) collected from Kāneʻohe Bay in a 400 mL Erlenmeyer flask. The mixture was shaken in a flask at 90 rpm in the dark for seven days. Following incubation, microplastics were removed by filtration using a 0.22 μm cellulose fiber filter, leaving a concentrated PVC leachate nominally equivalent to 1 g/L of PVC microplastics in seawater. This stock solution was then serially diluted with 0.22 μm FSW to produce the experimental treatments: 0.01 mg/L (low), 0.1 mg/L (moderate), and 1 mg/L (high) PVC leachate. Stock solution leachates were chemically characterized by GC-MS (full scan mode) and LC-MS/MS (MRM, multiple reaction monitoring mode) following the method of Saliu et al. (2020). These analyses indicated that the leachate contained phthalates at approximately 10% of the original PVC microplastic mass concentration. So while our treatments are characterized by the mass of PVC microplastic used to generate the leachate, the concentrations of chemical additives, such as phthalates, are estimated to be ~1 μg/L (low), ~10 μg/L (moderate), and ~100 μg/L (high).
Importantly, the selected exposure levels span reported environmental concentrations of phthalates in coastal surface waters, which range from 0 (limit of detection, LOD) to 168 μg/L (Mathieu and Bednarek 2022; Law et al. 1991; Hermabessiere et al. 2022; Paluselli et al. 2018; Net et al. 2015), addressing a common limitation of previous studies, which lack environmentally relevant exposure levels (Weis and Palmquist 2021). Our PVC leachate treatments reflect the full chemical mixture derived from UV-weathered PVC microplastics, rather than exposure to individual additives. Due to analytical limitations, exposure levels are reported as nominal values, consistent with established practices in leachate toxicity studies (Vered and Shenkar 2022; Tetu et al. 2019).
2.3 Imaging and annotation
Embryos preserved in 4% Z-Fix (aqueous zinc-buffered formalin diluted in 0.22 μm FSW) were permanently mounted on microscope slides using glycerol as a mounting medium. Imaging was conducted with a Nikon DS-Fi 3 camera attached to a Nikon Eclipse Ni-U manual microscope, and a composite image for each slide was generated through manual stitching. Images were analyzed using NIS-Elements BR software (version 5.02.01) to quantify the total number of embryos per treatment vial. We determined developmental stage reached of each embryo (egg, cleavage, morula, prawnchip, or early gastrula), and noted any morphological deformities or uncertainty due to fragmentation (typical, uncertain/torn, malformed). We chose to annotate the morphology of a torn embryo as uncertain, because coral embryos are known to continue developing even after fragmentation in early embryonic development (Heyward and Negri 2012). At 4 hours post-fertilization (hpf), samples are expected to reach the initial cleavage stage, characterized by the division of the fertilized oocyte into two blastomeres (Okubo et al. 2013). At 9 hpf, embryos are expected to reach the prawnchip phase, and at 14 hpf, coral embryos are expected to reach the early gastrula stage (Chille et al. 2022; Okubo et al. 2013). Examples of normally developed M. capitata embryos are shown in Figure 2. Our annotated images are available at https://github.com/sarahtanja/coral-embryo-scope/tree/main/scope-images.
2.4 Quantifying relative survival
At the beginning of the experiment, each vial contained two gamete bundles, corresponding to an average starting total of ~30 ± 6.4 eggs, based on previously reported estimates of 8-23 eggs per bundle (mean=15, SD=±5.1, SE=±0.3, [Padilla-Gamiño et al. (2011); Padilla-Gamiño and Gates (2012); ]). Viable embryos per vial at each time point were used as the response variable. Embryos classified as ‘typical’ or ‘torn’ were considered viable, while ‘malformed’ embryos were excluded from viable embryo counts. Torn embryos were included because early coral embryos can continue developing after fragmentation due to cellular totipotency (Heyward and Negri 2012). Eggs at 4 hpf were considered viable unless malformed; by 9 and 14 hpf any remaining eggs that were unfertilized or not developmentally progressing were classified as non-viable. Raw counts were imported and pre-processed in R (v4.5.1) using the tidyverse packages (Wickham et al. 2019). Treatment and time were coded as categorical factors and were explicitly ordered. Survival was analyzed as the counts of viable embryos at each time point. Distributions of viable embryo counts were visualized using boxplots with beeswarm overlays, showing individual vial-level observations across treatment × time combination (Figure 3). A two-way analysis of variance (ANOVA) was used to test the effects of treatment, developmental time (hpf), and their interaction on viable embryo counts. Model assumptions were evaluated using residual Q-Q plots, the Shapiro–Wilk test for normality, and Levene’s test for homogeneity of variances. All statistical tests were two-tailed with α = 0.05. Data analysis for embryo survival is available at https://github.com/sarahtanja/coral-embryo-scope/blob/main/code/03_survival.md.
2.5 Quantifying developmental timing
To assess if PVC leachate altered early embryonic developmental timing, we quantified the number of embryos in five developmental stages—egg, cleavage, morula, prawnchip, and early gastrula—for each sample. Because these data are multivariate counts and exhibited overdispersion (variance = 45.1, mean = 3.58), we used a multivariate generalized linear model (GLM) implemented in the mvabund package (mvabund_4.2.1, function manyglm; Wang et al. (2012) ). Counts of developmental stages were modeled simultaneously as a multivariate response (Y ~ treatment * hpf) using a negative binomial error distribution, with the log of the embryo counts in each sample as an offset term. The offset term ensures that the model captures shifts in the ratios of stages rather than in absolute counts for each developmental stage. The full dataset was analyzed as a single multivariate response matrix, allowing us to detect both overall shifts in development and stage-specific responses. Model significance was assessed using PIT-trap resampling with 999 iterations, which provides robust p-value estimation under non-normal residual structure. We report multivariate test statistics (evaluating changes in developmental stage across treatments and time) and univariate tests for each stage. Significance of main effects and interaction terms was assessed using Analysis of Deviance (Type I tests). All analyses were performed in R (v4.5.1) with α = 0.05. The R code used for the developmental timing analysis is available at https://github.com/sarahtanja/coral-embryo-scope/blob/main/code/04_timing.md .
2.6 Quantifying morphological abnormality
Similarly to developmental timing, we analyzed morphological status using a multivariate generalized linear model (GLM). We tested whether PVC leachate altered the counts of embryos classified as typical, uncertain (torn), or malformed across developmental time. For each sample, embryo counts were grouped into these three categories. Because the data were overdispersed (variance = 76.5, mean = 5.97), we specified a negative binomial error distribution. To account for changes in embryo counts due to survival, we included the log of the embryo counts in each sample as an offset term. The multivariate response matrix (Y) included counts of typical, uncertain (torn), and malformed embryos. The model tested the effects of treatment, developmental time, and their interaction (Y ~ treatment * hpf). The R code and our analysis for morphological abnormality are available at https://github.com/sarahtanja/coral-embryo-scope/blob/main/code/05_abnormality.md.
2.7 DNA/RNA extractions and quantification
At each developmental endpoint (4, 9 and 14 hpf), coral embryos were transferred from exposure vials to 2 mL microcentrifuge tubes, preserved in DNA/RNA ShieldTM (Zymo Research, USA), and stored at -80°C. DNA and RNA were extracted simultaneously from the same pooled samples using the Quick-DNA/RNATM Miniprep Plus Kit (Zymo Research, USA). Samples of embryos in DNA/RNA ShieldTM were thawed to room temperature and pooled with up to three coral colony crosses from the same treatment group to achieve sufficient DNA/RNA yield (pooled sample structure is shown in Figure 16). Samples were pooled in ZR BashingBead Lysis TubesTM (0.1 and 0.5 mm) (Zymo Research, USA) and homogenized for 10 minutes using a Mortexer. Extraction and purification steps followed the manufacturer’s cell preparation protocol, but skipped the proteinase-K digestion step. Purified total RNA was quantified using a Qubit-4 Fluorometer with the RNA High Sensitivity Assay Kit (ThermoFisher, USA) and shipped on dry ice to Azenta Life Sciences for library preparation with Poly(A) selection. DNA was similarly quantified using the dsDNA Broad Range (BR) Assay Kit (ThermoFisher, USA) according to the manufacturer’s protocol. This resulted in 63 pooled samples from which both DNA and RNA were extracted. These samples are paired, i.e. DNA and RNA was extracted from the same embryos for each of the 63 pooled samples. Each pooled sample represents genetic variation from up to six parent colonies (Figure 16).
2.8 Transcriptomics (RNA library prep, sequencing, processing, and analysis)
2.8.1 RNA library preparation, and sequencing
Our 63 pooled samples of total purified RNA were processed for library preparation and sequencing (Azenta Life Sciences). Libraries were prepared using Poly(A) selection to target eukaryotic mRNA, with a sequencing depth of 20 million paired-end reads per sample. Sequencing was performed on an Illumina NovaSeq X+ platform (x150bp). Raw paired-end reads (FASTQ format) were submitted to the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) under BioProject accession number PRJNA1177827.
2.8.2 Quality checks, alignment, and gene count matrix generation
Raw paired-end FASTQ files were checked for quality with FastQC (v0.12.1) and summarized using MultiQC (v1.14) (Ewels et al. 2016). Adapter sequences were trimmed, and sequences were pre-processed with fastp (v0.23.2) (Chen 2023)using automatic adapter detection for paired-end data, a phred quality score threshold of 30, removal of polyX tails ≥6 bp, and fixed trimming of 10 bp from the 5′ end of both forward and reverse reads. Trimmed reads were aligned to the M. capitata reference genome (HIv3; Rutgers Coral Genome Project (Stephens et al. 2022)) using HISAT2 (v2.2.1) (Kim et al. 2019) with splice-aware alignment. Exon coordinates and splice junctions were extracted from the genome annotation file (GTF) with HISAT2 helper scripts provided during alignment to improve mapping across intron–exon boundaries. The resulting SAM files were converted to sorted and indexed BAM files using SAMtools (v1.12) (Danecek et al. 2021). Transcript assembly and abundance estimation were performed with StringTie (v2.2.1)(Pertea et al. 2015), guided by the reference annotation (GFF3). A gene count matrix (genes x samples) was generated using the StringTie helper script prepDE.py3, enabling downstream differential expression analysis. The quality-assurance, quality-check (QAQC) and alignment R code can be viewed at https://github.com/sarahtanja/coral-embryo-RNAseq/tree/main/code.
2.8.3 Differential gene expression analysis with DESeq2
Raw gene counts were imported into R (v4.2.3) along with sample metadata. Genes with fewer than 10 counts in at least 85% of samples were removed prior to analysis. Count data were normalized using DESeq2 (v1.38.3) (Love et al. 2014), with size factors estimated and variance-stabilizing transformation (VST) applied for downstream visualization. Principal component analysis (PCA) was performed on the 500 most variable genes to assess global sample structure and identify potential outliers (Figure 6 A). Outlier samples were flagged by visual inspection and Z-score thresholding (>3 standard deviations along PC1 or PC2) and subsequently removed. Two samples were excluded due to poor alignment in HISAT2 (131415L4 0.2%, 789C4 8.9%; MultiQC Bowtie2/Hisat2 report) and failure to cluster with other samples in the PCA plot. The remaining 61 samples had alignment rates ranging from 27.4% to 83.2%. Following outlier removal, gene filtering was repeated using the same filtering thresholds (retaining genes with ≥ 10 counts in at least 85% of samples). Sample-to-sample relationships were further analyzed using a z-score transformed heatmap with hierarchical clustering, with samples annotated by spawn night, leachate treatment and developmental stage (Figure 6 B).
Our DESeq2 (v1.38.3) (Love et al. 2014) model accounted for spawn night, stage, leachate, and the interaction between leachate and stage (~ spawn_night + stage + leachate + stage:leachate). We chose to treat leachate and hours post fertilization as categorical factors to test for stage-specific differences without assuming a particular dose-response shape, since we were looking for non-linear response trends. To evaluate model structure, we conducted likelihood ratio tests (LRTs). We first analyzed the effect of spawn night by comparing the full model to a reduced model excluding spawn night. This analysis identified 3,551 genes (~29%) as significantly differentially expressed (Benjamini-Hochberg (BH) FDR-adjusted p-value < 0.05) indicating that spawn night contributes substantially to expression variability and represents a batch effect. Accordingly, spawn night was retained in the full model to control for the variability in gene expression due to the different nights on which our parent coral colonies spawned. We then used LRTs to confirm the inclusion of both leachate and the stage x leachate interaction terms. Following these tests, Wald contrasts were performed to estimate log2 fold changes for pairwise contrasts, specifically assessing the effect of each leachate level at each developmental stage relative to the control, as well as the larger signal across developmental stages. Genes with false discovery rate (FDR) adjusted p < 0.05 were considered significantly differentially expressed in the LRT tests and Wald tests. We generated lists of DEGs responsive to leachate and to developmental progression.
We then grouped leachate-responsive DEGs and developmental progression DEGs by expression patterns. Because the number of leachate-responsive DEGs was relatively small (n=130) and pre-filtered, the scale-free topology criteria were not met; therefore expression patterns were summarized using trajectory-based clustering implemented in DEGreport v1.46.0, rather than co-expression network analysis. In contrast, the larger set of development-related DEGs (10,793 genes) was analyzed using weighted gene co-expression network analysis (WGCNA) with the WGCNA R package (v 1.73)(Langfelder and Horvath 2008).
2.8.4 Gene ontology (GO) enrichment and slim characterization
We characterized the GO terms associated with all of our DEGs for each expression pattern subset. Significant genes identified in the DESeq2 analysis were functionally annotated and assessed for Gene Ontology (GO) enrichment. Protein-coding sequences from the Montipora capitata genome v3 (http://cyanophora.rutgers.edu/ montipora/) were matched to EggNOG annotations, and gene lengths were extracted from the reference GFF3 file to account for length-dependent bias in RNA-seq detection. GO enrichment analysis was conducted using the R package goseq (v1.50.0) with Wallenius’ distribution to correct for gene length bias (the increased detection power for longer genes) and restricting tests to biological process terms. A background universe of the filtered DESeq2 gene set (12,221 genes) was used. Significance was determined by Benjamini–Hochberg false discovery rate (FDR) correction, with GO terms retained at adjusted p< 0.05.
2.9 Microbiome (DNA library prep, sequencing, processing, and analysis)
2.9.1 DNA Amplification, Library Preparation, and Sequencing
Purified DNA for each of our 63 pooled samples was used directly for library preparation, without fragmentation or size selection. Amplicon libraries targeting the V3–V4 region of the 16S rRNA gene were generated following Illumina’s 16S Metagenomic Sequencing Library Preparation protocol (Illumina, Part no. 15044223 Rev. B) using primers 5′-CCTACGGGNGGCWGCAG-3′ and 5′-GACTACHVGGGTATCTAATCC-3′ (Klindworth et al. 2013). PCR amplification was performed according to the manufacturer’s guidelines. Libraries were sequenced at the University of Washington Microbiome Initiative (mim_c) on an Illumina NextSeq™ 2000 platform using the P1 600-cycle reagent kit. Raw paired-end reads were deposited in the NCBI Sequence Read Archive under BioProject accession number PRJNA1244351.
2.9.2 Amplicon Processing and Taxonomic Inference
All sequence pre-processing steps were performed in QIIME2 v2023.9 (Bolyen et al. 2019). Demultiplexed paired-end reads were trimmed with Cutadapt to remove primer sequences and then denoised with DADA2. QIIME2 was used to examine rarefaction curves, assess sequencing depth, infer amplicon sequence variants (ASVs), remove chimeras, and construct a feature table. Representative ASVs were taxonomically assigned using a pre-trained SILVA Naive Bayes classifier. ASVs classified as mitochondria, chloroplast, or unassigned were removed from the filtered feature table prior to downstream analyses.
2.10 Microbial community compositional data analysis
Data representing our 63 pooled samples coral embryo microbiome communities. Amplicon sequence variant (ASV) count data were analyzed following recommendations for microbiome sequence data, which are constrained by relative abundance and sequencing depth rather than absolute counts (Gloor et al. 2017). Analyses were conducted in R (v4.5.1) using the packages tidyverse, qiime2R, phyloseq, vegan, compositions, and zCompositions.
Because log-ratio transformations cannot accommodate zeros directly, zero counts were first replaced using count zero multiplicative replacement (cmultRepl, method = “CZM”). The zero-adjusted count matrix was then transformed using the centered log-ratio (CLR) transformation to map compositional data into Euclidean space suitable for standard multivariate analyses. The resulting CLR matrix contained 63 samples and 663 retained ASVs.
Between-sample dissimilarity (Beta diversity) was quantified using Aitchison distance, calculated as Euclidean distance on the CLR-transformed feature table. Principal components analysis (PCA) was then performed on the CLR-transformed matrix using the prcomp() function. To evaluate whether differences among groups reflected unequal within-group variability rather than centroid separation, homogeneity of multivariate dispersion was tested using betadisper(). Dispersion was evaluated separately for spawn night, leachate treatment, developmental stage, and the leachate × stage interaction.
Differences in microbial community composition across experimental factors (∼leachate*stage) were tested using permutational multivariate analysis of variance (PERMANOVA) implemented with adonis2() in the vegan package, with 999 permutations. To account for non-independence among samples collected on the same spawning night, permutations were constrained within spawn night using the strata argument. Terms were tested sequentially using the by = “terms” option. This approach allowed effects of leachate exposure and developmental timing to be evaluated while controlling for batch effects associated with the night of spawning.
2.10.1 Differential abundance & prevalence testing with MaAsLin3
Associations between microbial features and experimental variables were tested using MaAsLin3 (Mallick et al. 2021) in R. Analyses were performed on taxonomic features collapsed to the genus level (Level 6). For both data types, features were retained if present in ≥5% of samples with relative abundance ≥0.1%. No additional normalization was applied, as inputs were proportion-normalized during preprocessing. Differential abundance and prevalence testing were performed using a generalized linear mixed-effects modeling framework implemented in MaAsLin3 with the formula: abundance/prevalence ~ leachate * hpf + (1|spawn_night). Significance was determined by adjusted joint q-values < 0.1, which were corrected for multiple testing with the Benjamini–Hochberg method for adjusting for false discovery rate.
2.10.2 Functional Prediction with PICRUSt2
Predicted functional profiles were inferred from 16S rRNA amplicon sequence variants (ASVs) using PICRUSt2 (Douglas et al. 2020). A total of 547 PICRUSt2 predicted MetaCyc pathways were included in downstream modeling. Differential abundance and prevalence testing of MetaCyc predicted pathways was conducted using MaAsLin3 in R.
3 Results
3.1 H1: Survival and Development
3.1.1 Survival
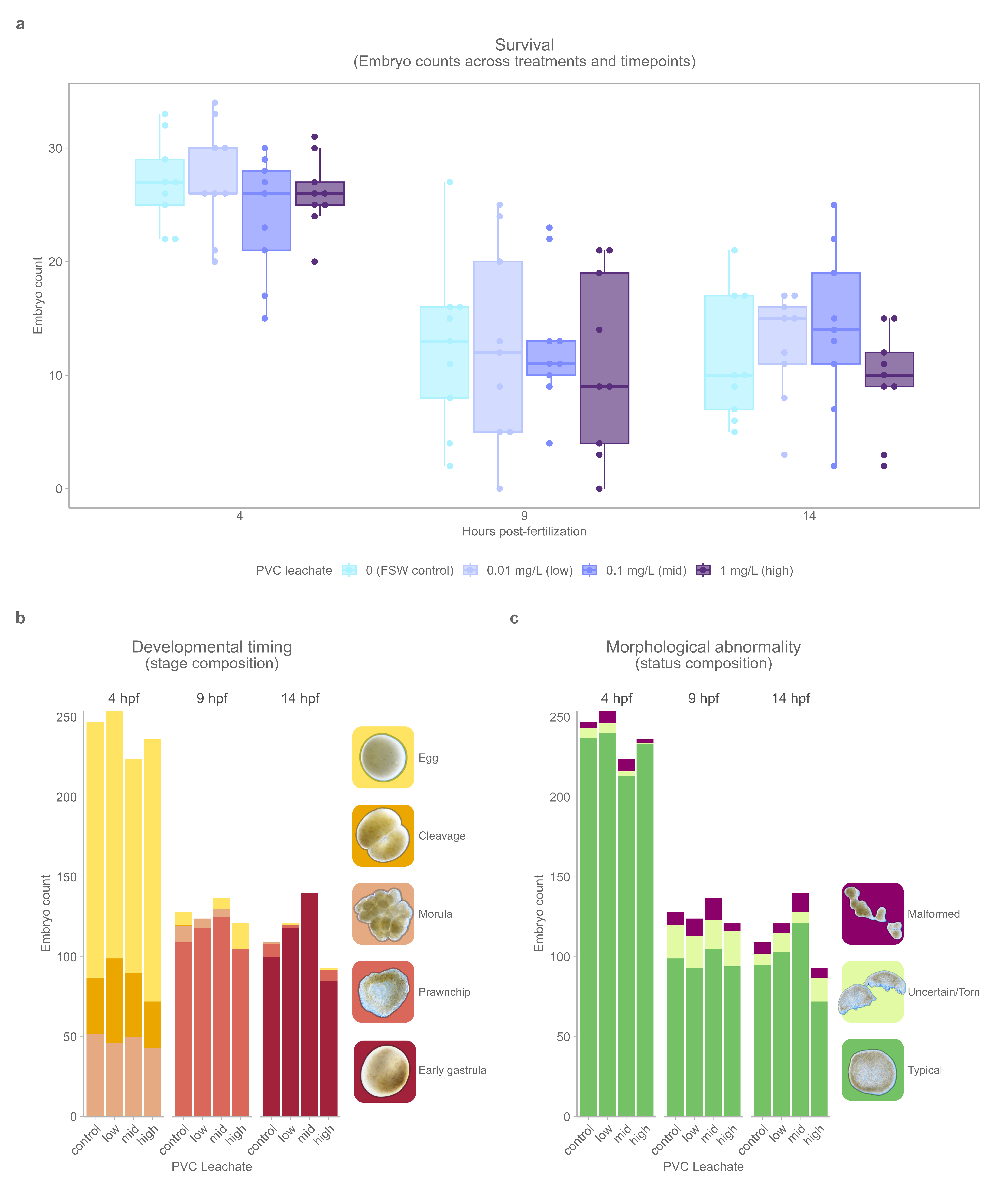
Polyvinyl chloride (PVC) leachate did not change embryo survival during the first 14 hours post-fertilization (hpf) of embryonic development (Figure 3 a; treatment: F3,96 = 0.523, p = 0.668)(Table 2).
Time was the main factor affecting embryo survival. Mean counts of surviving embryos across replicate vials declined from 26.1 ± 4.4 SD at 4 hpf to 12.2 ± 7.4 SD at 9 hpf, and then remained relatively stable between 9 and 14 hpf (11.9 ± 5.7 SD) (hpf: F2,96 = 64.04, p < 0.0001). Survival was similar across all PVC leachate treatments (interaction: F6,96 = 0.572, p = 0.752), showing that neither exposure level nor exposure duration altered embryo survival trends. Model assumptions for our two-way ANOVA were met as residuals were normally distributed (Shapiro–Wilk normality test W = 0.99, p = 0.69) and variances were similar among groups (Levene’s F~11, 96~ = 1.25, p= 0.26 ).
3.1.2 Morphological abnormality
Morphological abnormality (embryos annotated as typical, uncertain/torn, or malformed) did not vary across PVC leachate treatments. The multivariate negative binomial model found that neither PVC leachate treatment (treatment: Dev = 4.67, p = 0.607) nor the relationship between PVC leachate treatment and developmental time (interaction: Dev = 11.41, p = 0.770) had any effect on morphological abnormality ( Table 4).
However, we found developmental stage significantly impacted morphology status. Regardless of treatment, we found an increase in uncertain embryos (Dev = 25.24, p = 0.002) at the 9 hpf (prawnchip) stage, with a concurrent decrease in typical embryos (Dev = 63.75, p = 0.001)( Figure 3 C). Malformed embryos however did not vary significantly across developmental time (Dev = 1.91, p = 0.366) (Table 4).
Mean count summaries show that typical embryos were most abundant at 4 hpf across all treatments, then declined at 9 and 14 hpf, while uncertain embryos tended to peak at 9 hpf. Malformed embryos remained consistently low. Together, these results indicate that PVC leachate exposure had no significant effect on the counts of embryos classified as typical, uncertain/torn, or malformed (Figure 3 c). Overall the number of embryos found to be malformed were low compared to typically-formed embryos.
Anecdotaly, we observed that the secondary cell division (progressing from the 2-cell to the 4-cell stage) initiates at the center of the two blastomeres and extends outward (Figure 2 G). This contrasts with the primary cell division (initial cleavage) which begins at one pole of the fertilized egg and progresses across, which is evident in the heart-shaped embryos (Figure 2 C).
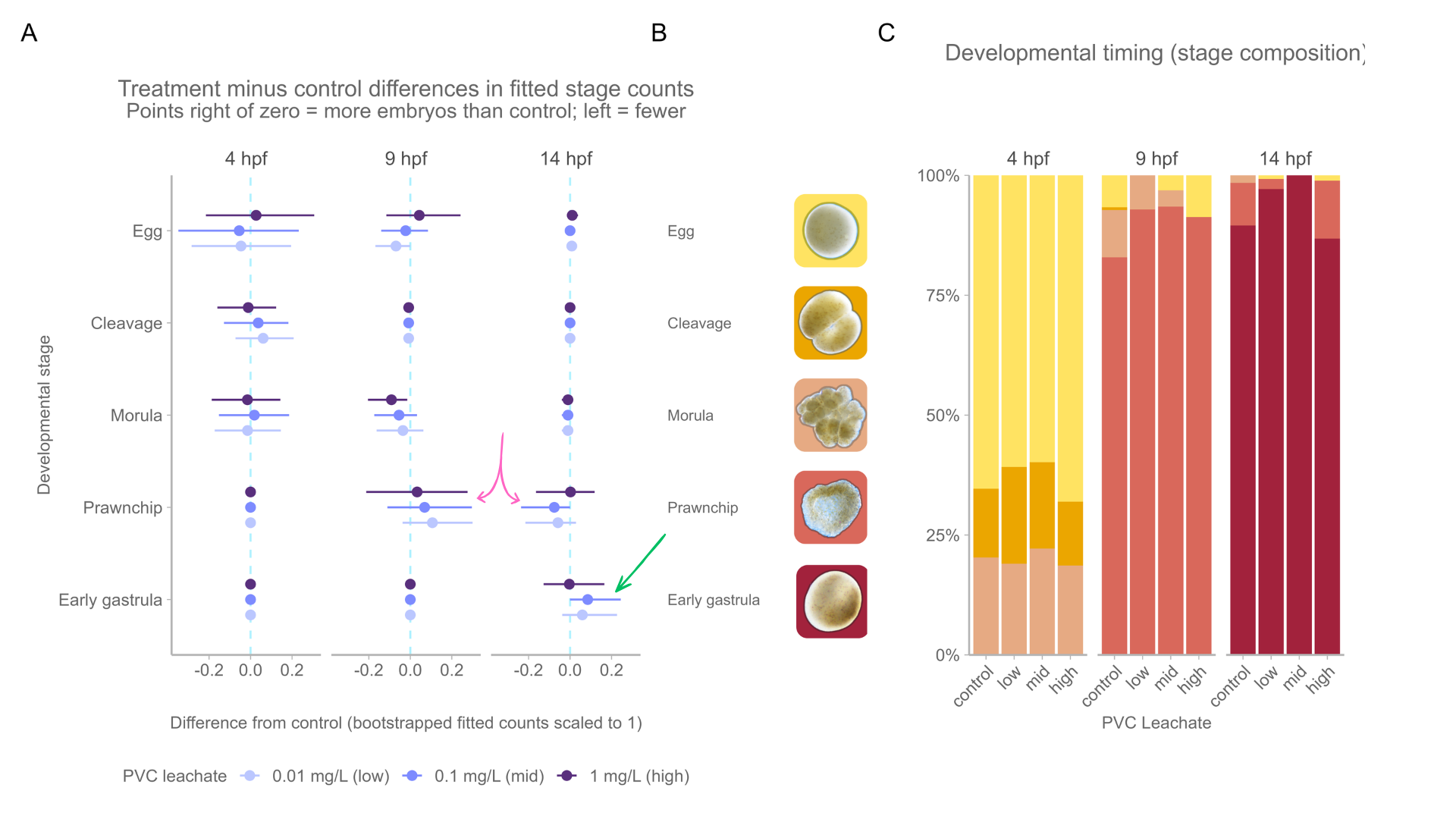
3.1.3 Developmental timing
We tested whether the mix of embryo developmental stages changed over time across PVC leachate treatments using a multivariate generalized linear model. As expected, embryo stage composition changed strongly with time (hpf: Dev = 669.0, p < 2×10-16 ). At 4 hpf, most embryos were in the egg, cleavage, or morula stages. At 9 hpf, most embryos were in the prawnchip stage. By 14 hpf, the early gastrula stage dominated (Figure 3 B).
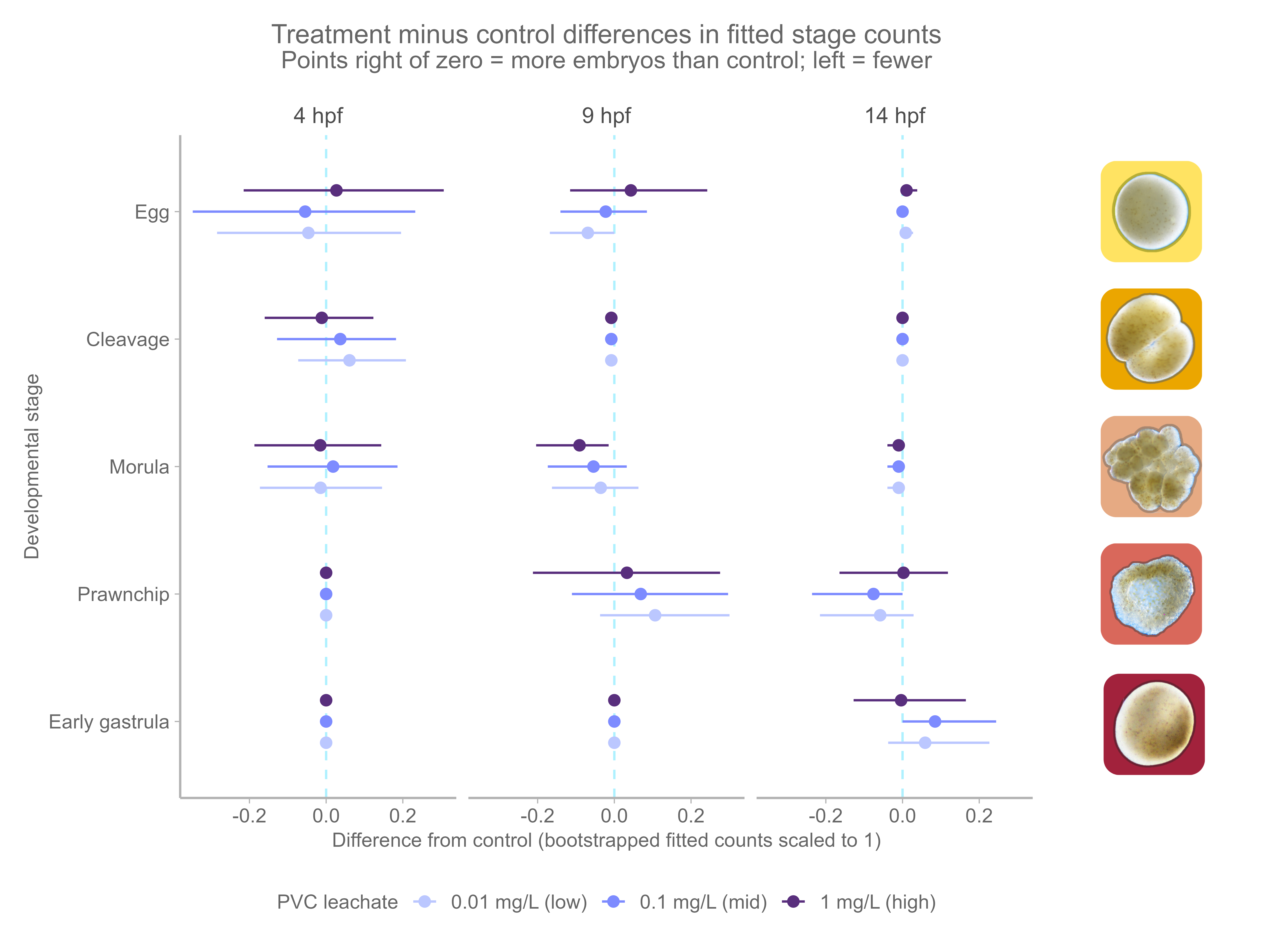
PVC leachate exposure alone did not cause an overall shift in the stage composition of embryos (treatment: Dev = 1.2, p = 0.999). However, treatment effects changed across time (interaction: Dev = 48.1, p = 0.002). When stages were examined separately, we found this pattern was driven mainly by differences in the prawnchip stage at 14 hpf (the interaction effect is localized to a specific prawnchip stage developmental window), with weaker trends in the egg and morula stages. No treatment effects were detected in the cleavage or early gastrula stages.
Overall, these results suggest that PVC leachate did not broadly disrupt normal developmental timing, but it may have caused small, stage-specific changes, possibly speeding the morula to prawnchip transition at 9 hpf, and the prawnchip to early gastrula transition at 14 hpf under low and mid, but not high leachate (Figure 4, OR Figure 5).
3.2 H2: Gene expression
3.2.1 Global expression
Principal component analysis (PCA) was performed on variance-stabilized gene counts using the top 500 most variable genes as an exploratory tool to assess global patterns of variation. The first two principal components accounted for 72.4% of the total variance among samples and revealed strong clustering by early embryonic stage, with separation primarily along principal component 1 (PC1, 53.9% of global sample structure variance) (Figure 6 A). Samples formed three distinct, non-overlapping clusters with minimal within-group dispersion, indicating that stage represents the dominant source of variance in the dataset. Principal component 2 (PC2, 18.5% of global sample structure variance) primarily captures the variation within groups, indicating that secondary sources of variation are present but do not override the dominant stage effect. Treatment-associated effects of PVC leachate were not readily apparent in the leading principal components. The pronounced stage-driven structure is consistent with large-scale transcriptional reprogramming during early embryogenesis ( Figure 6 B).
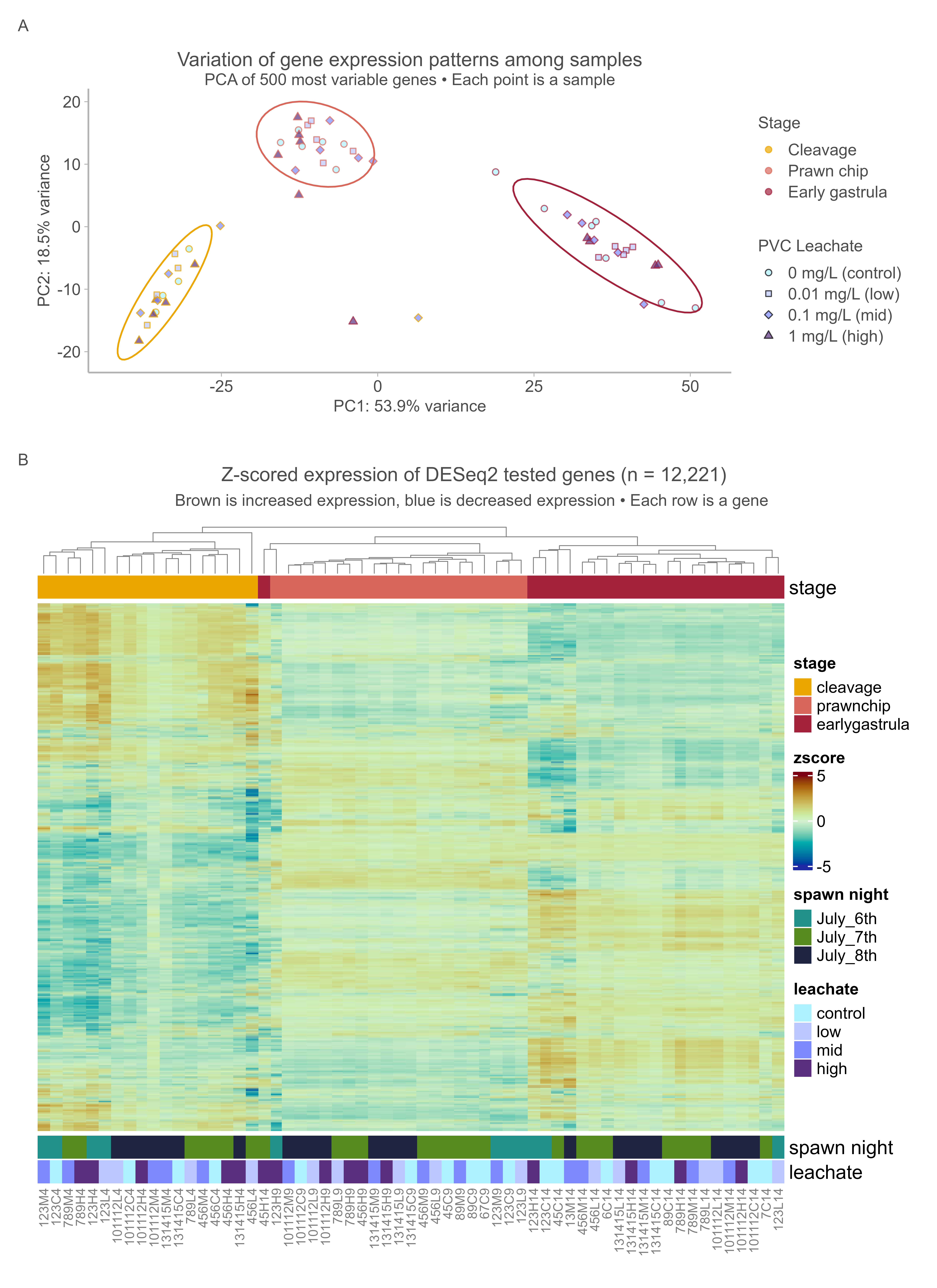
We followed visual PCA exploration with a permutational multivariate analysis of variance (PERMANOVA) to statistically test for treatment-associated effects. A PERMANOVA with 999 permutations was performed on the Euclidean distance matrix of transformed gene expression values to assess the effects of leachate level, developmental stage, and their interaction on global expression profiles while controlling for variation among spawning nights (permutations were restricted or ‘stratified’ within spawn night by setting the strata = spawn_night). The PERMANOVA identified developmental stage as the dominant driver of multivariate expression differences, explaining ~59% of the overall variance in gene expression ( R² = 0.59, F = 41.22, p = 0.001 ), reflecting our strong stage-specific transcriptional patterns. In contrast, neither leachate level ( R² = 0.02, F = 1.09, p = 0.338 ) nor the interaction between leachate and stage ( R² = 0.04, F = 0.81, p = 0.679 ) significantly influenced the multivariate expression structure. Residual variation accounted for 35% of the total variance. Tests for homogeneity of multivariate dispersion revealed significant within-stage differences (F = 15.03, p = 0.001, 999 permutations), suggesting that the observed PERMANOVA stage effect reflects a combination of strong centroid separation and variable spread. Despite differences in dispersion, the magnitude of the stage effect and the clear separation observed in PCA indicate that developmental stage represents robust, biologically meaningful early developmental gene expression programs.
3.2.2 DEG clusters
There are ~56K genes in the M. capitata genome (Stephens et al. 2022), of which we filtered to a test group of 12,221 genes. Of the 12,221 genes tested with DESeq2, we found 130 were differentially expressed due to the interaction between leachate and stage. We will refer to these as the leachate-responsive DEGs.
Among the 130 leachate-responsive DEGs, trajectory-based clustering using the DEGreport v 1.44.0 R package and the degPatterns function found 128 genes that fit into one of three gene expression patterns (Figure 7). These expression patterns are structured by developmental timing, with groups exhibiting highest relative transcript abundance at cleavage (31 genes), prawn-chip (25 genes), or early gastrula (75 genes) stages, respectively (Figure 7 A-C). Two leachate-responsive DEGs did not fit into any of these patterns, and were excluded from downstream analysis. Leachate effects within these clusters were modest, with leachate treatments exhibiting subtle deviations compared to the control samples most evident at stages where baseline expression was lowest.
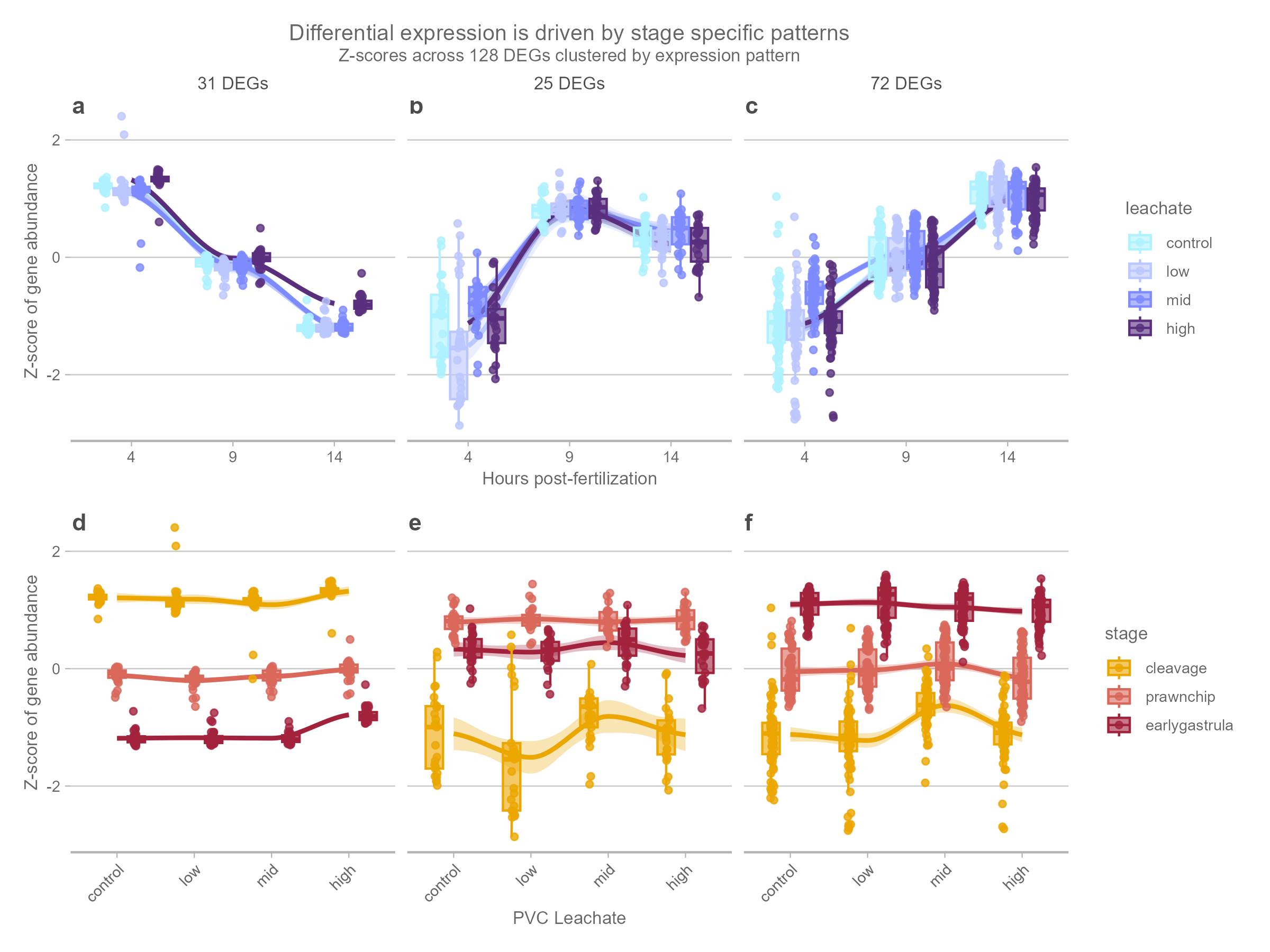
In contrast, the DEGs associated with development (10,793 genes) were analyzed using weighted gene co-expression network analysis (WGCNA) with the WGCNA v 1.73 R package (Langfelder and Horvath 2008), which identified six modules ranging in size from 326 to 2,805 genes. Module eigengene profiles revealed four dominant developmental programs (expression patterns): a cleavage-high program characterized by decreases in expression over developmental time, two prawn-chip–associated programs showing either up-regulation (hump or inverted-U-shaped expression profile) or mid-stage suppression (valley or U-shaped expression profile), and late developmental programs with increasing expression toward early gastrulation. These modules represent coordinated transcriptional programs associated with embryonic progression and were retained for downstream GO functional enrichment analyses. 149 developmental DEGs were not assigned to a cluster in the WGCNA, and were excluded from downstream analysis.
All 130 leachate-responsive DEGs are also listed in the genes that change across developmental stage; indicating that the shifts in gene expression detected across leachate treatments are nested within the larger shifts in gene expression occurring over developmental time.
3.2.3 Summary of GOslim terms
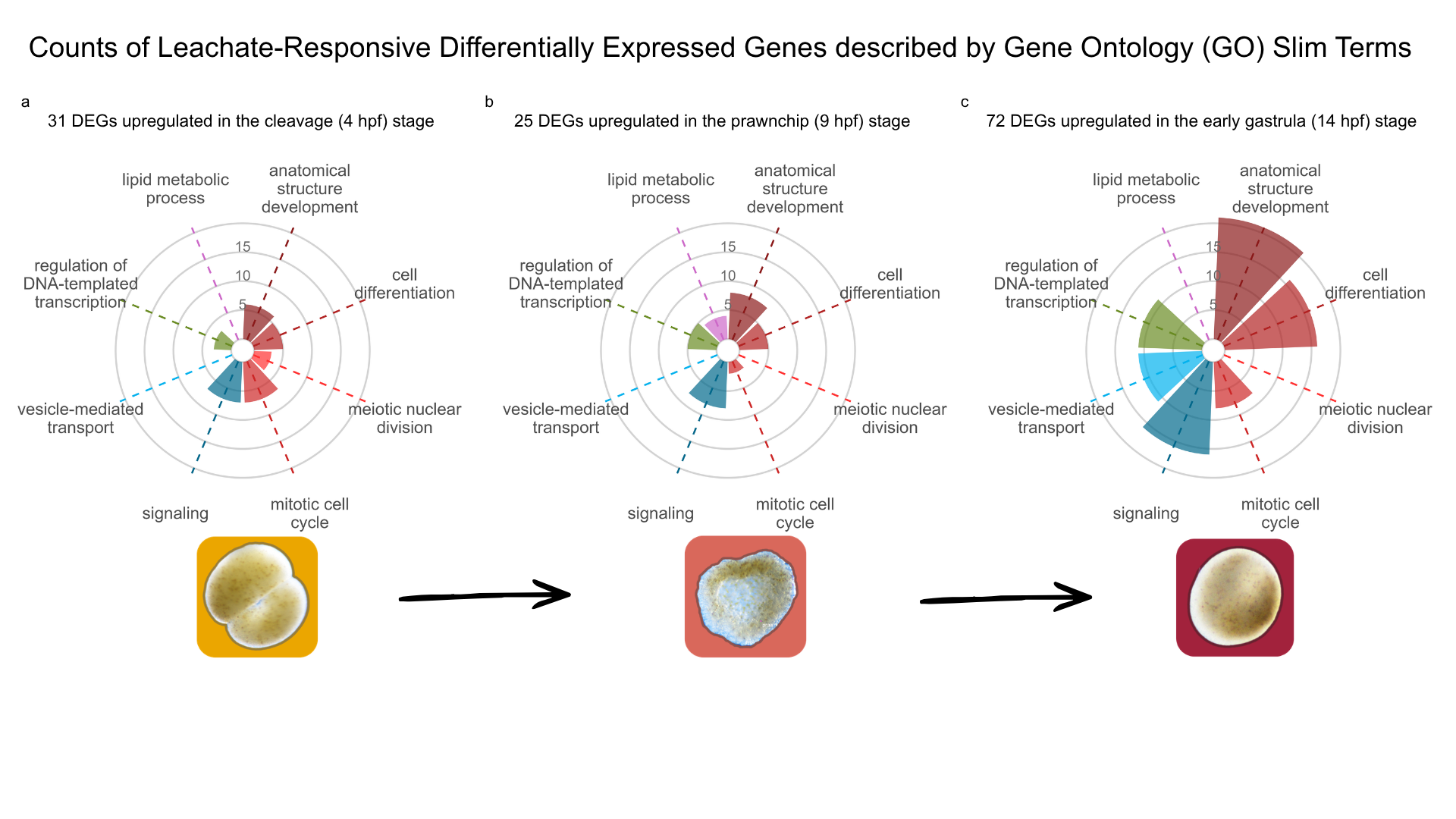
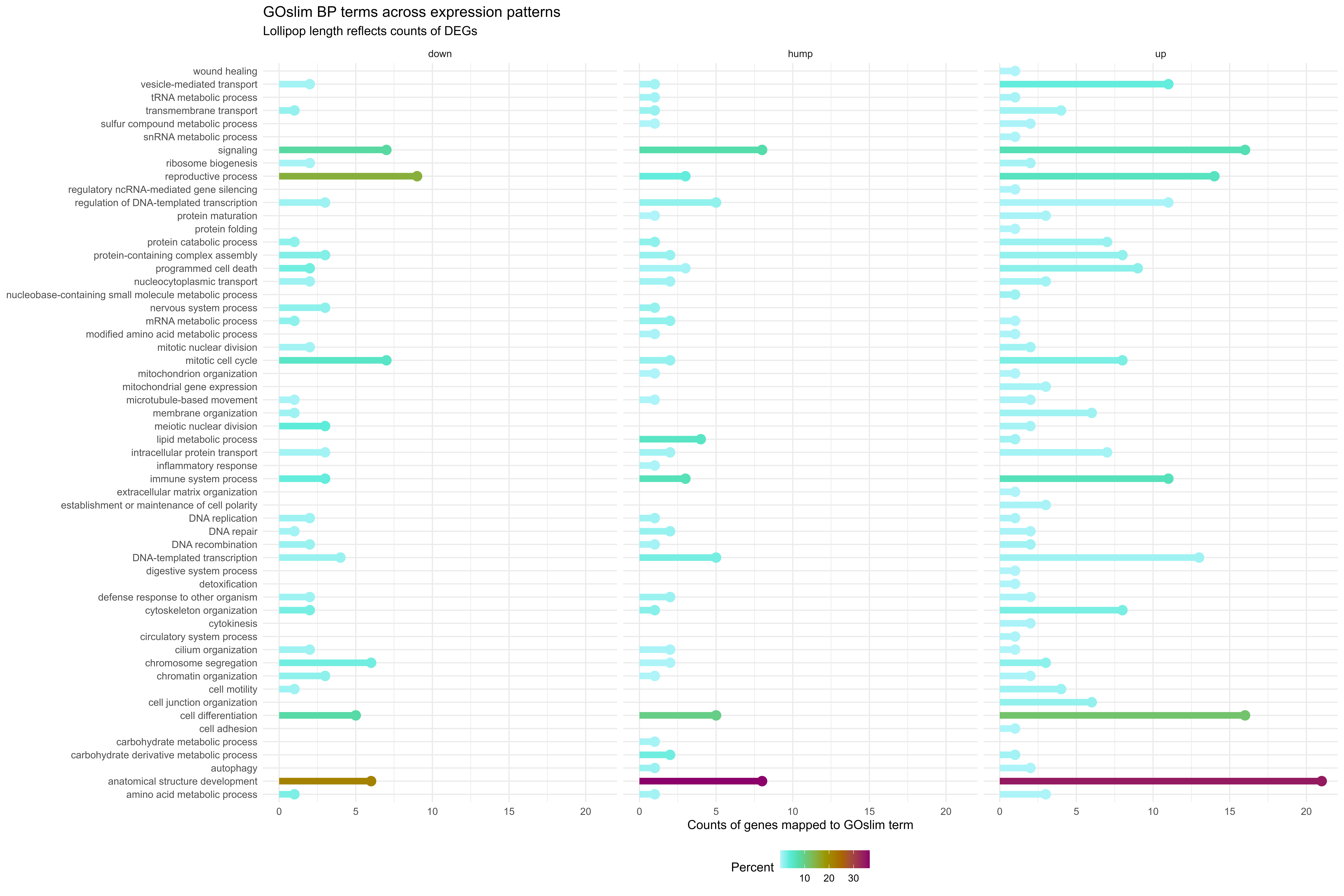
Of the 12,221 genes included in the DESeq2 testing universe, 6,846 (56%) had at least one Gene Ontology (GO) annotation. GO annotation coverage among leachate-responsive differentially expressed genes (DEGs) was high across all stage-associated expression patterns, including 27/31 (87%) genes in the cleavage-associated (4 hpf) pattern, 19/25 (76%) genes in the prawnchip-associated (9 hpf) pattern, and 62/72 (86%) genes in the early gastrula-associated (14 hpf) pattern. GOseq enrichment analysis did not identify any significantly enriched biological processes after correction for gene-length bias (q > 0.05 for all comparisons). Therefore, GO Slim categories were used descriptively to summarize the functional identities of leachate-responsive genes within each developmental expression pattern (Figure 8).
The 31 leachate-responsive DEGs exhibiting the cleavage (4 hpf) expression pattern, in which transcript abundance decreased across development (4>9>14 hpf), were dominated by GO Slim terms related to early developmental regulation and nuclear organization, including anatomical structure development, cell differentiation, meiotic nuclear division, chromosome segregation, and chromatin organization. Additional cleavage-associated terms included intracellular protein transport and nervous system process. Together, these functions suggest that leachate-responsive genes expressed most strongly during cleavage were associated with maternally provisioned developmental programs, chromosomal organization, and cell division fidelity during early embryogenesis.
In contrast, the prawnchip expression pattern, characterized by genes peaking transiently at 9 hpf (4<9>14 hpf), contained fewer total leachate-responsive DEGs (25) and was characterized by genes involved in lipid metabolic process, signaling, and programmed cell death. These functions suggest a transient shift toward metabolic and stress-associated responses during the prawnchip stage.
The early gastrula (14 hpf) expression pattern, in which transcript abundance increased across development (4<9<14 hpf), contained the largest number of leachate-responsive DEGs (72 genes). Dominant GO Slim categories included anatomical structure development, cell differentiation, signaling, vesicle-mediated transport, mitotic cell cycle, cytoskeleton organization, and protein-containing complex assembly (). These functions are consistent with increased cellular remodeling, intracellular trafficking, and structural reorganization during gastrulation.
Several GO Slim categories, including anatomical structure development, cell differentiation, DNA-templated transcription, regulation of DNA-templated transcription, reproductive process, immune system process, and signaling, were consistently observed across all three developmental expression patterns, suggesting that core developmental and regulatory processes were responsive to PVC leachate exposure throughout embryogenesis. However, stage-specific differences in the dominant functional categories indicate that the biological consequences of leachate exposure varied across developmental time, with cleavage-stage responses emphasizing chromosomal and nuclear regulation, prawnchip responses reflecting transient metabolic and stress-associated processes, and early gastrula responses dominated by cellular activation and structural reorganization.
3.3 H3: Microbiome
3.3.1 Alpha diversity
A two-way analysis of variance (ANOVA) revealed no significant effects of PVC leachate level, developmental stage, or their interaction on Shannon diversity. Differences in Shannon diversity among leachate treatments were not statistically significant (F₃,₄₈ = 1.25, p= 0.302), nor were differences among developmental stages (F₂,₄₈ = 1.11, p= 0.339). Similarly, the leachate × stage interaction was not significant (F₆,₄₈ = 0.33, p= 0.919), indicating that leachate did not affect microbial alpha diversity across stages. A two-way ANOVA revealed a significant effect of developmental stage on Faith’s phylogenetic diversity, with post hoc Tukey HSD tests showing reduced phylogenetic diversity at 14 hpf compared to 4 hpf (padj = 0.04).
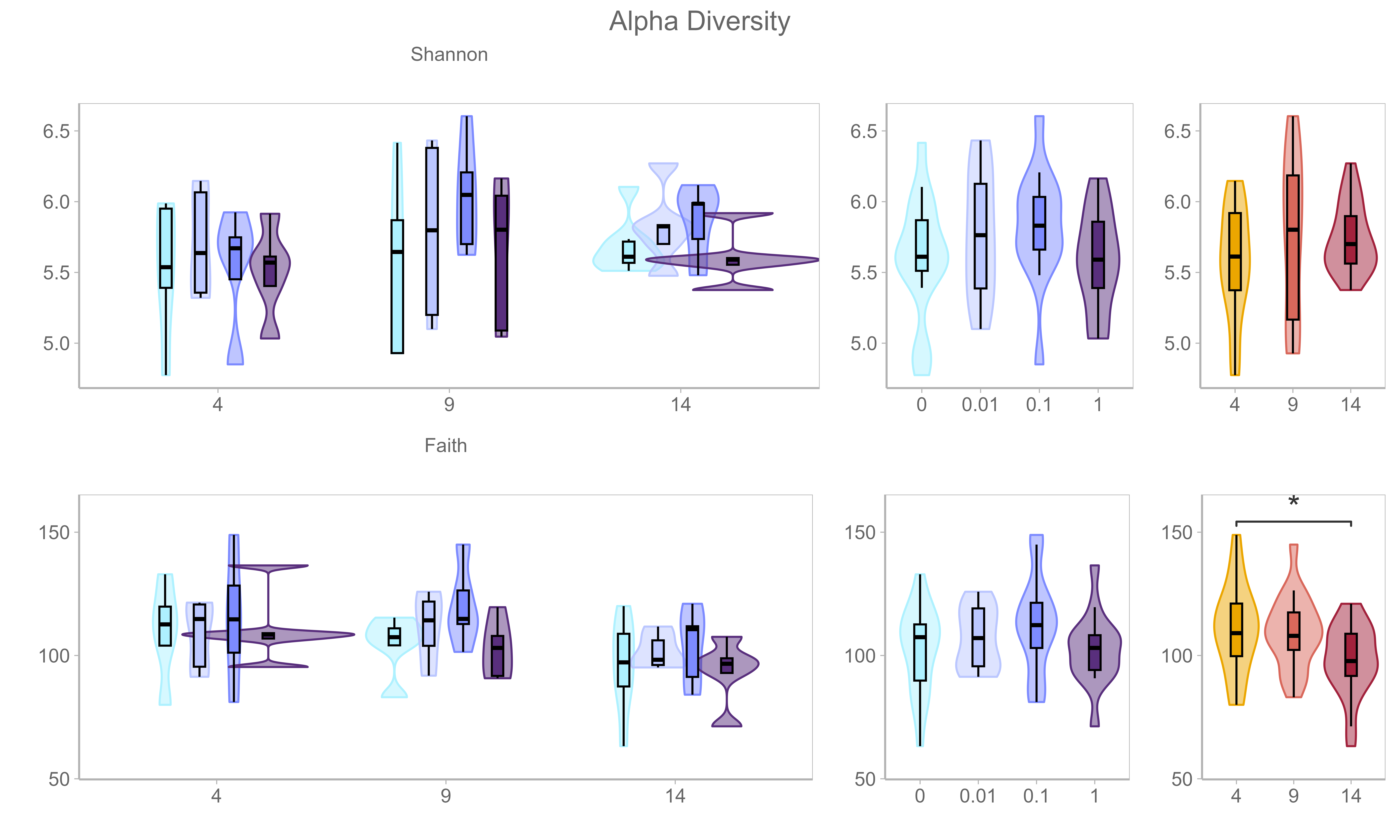
Alpha diversity remained stable across treatments, indicating limited sensitivity of within- sample microbial diversity to leachate exposure or developmental progression.
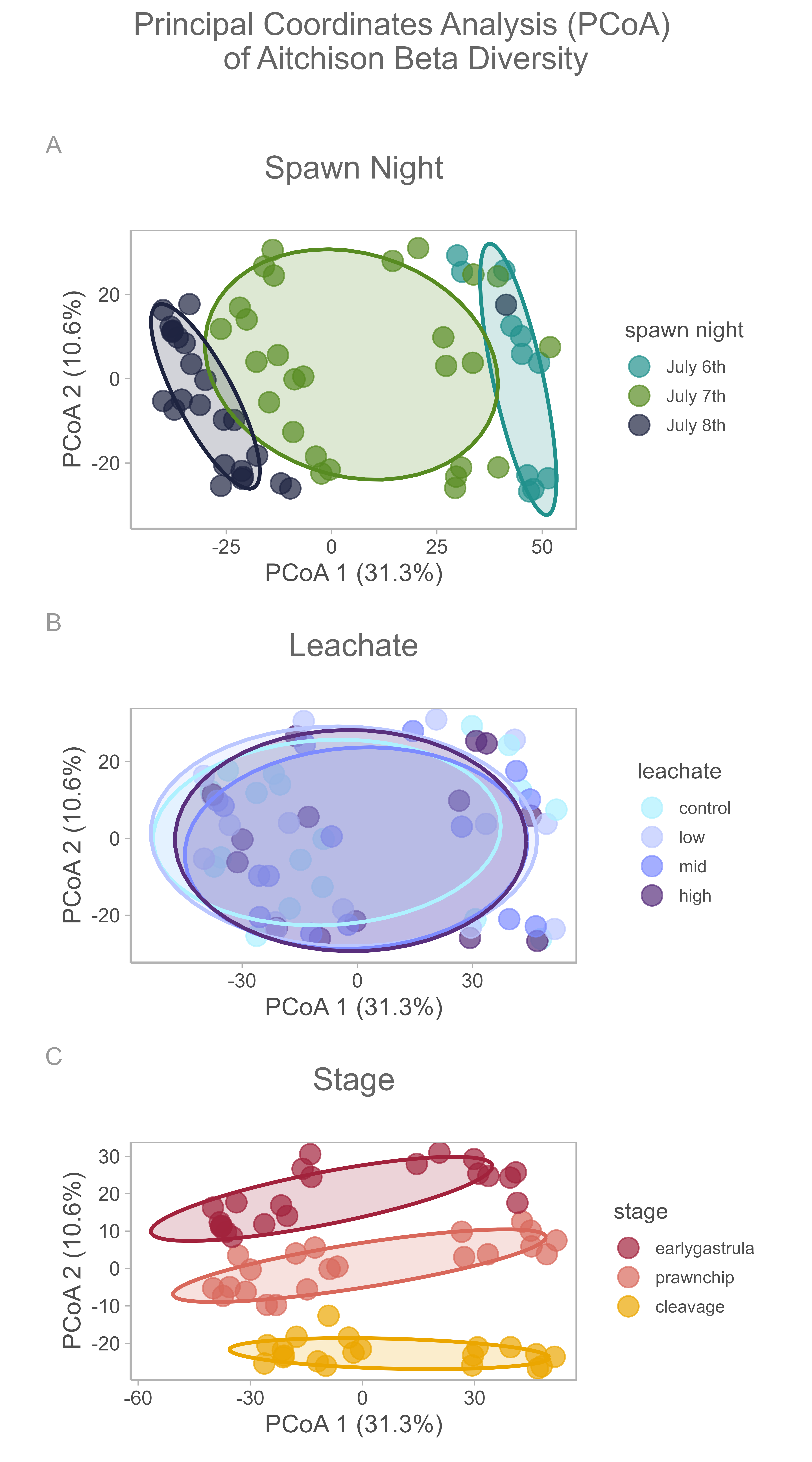
Following recommendations from Gloor et al. (2017), we used the centered log-ratio (CLR) transformation to convert our data to Euclidean space to an Aitchison distance (Euclidean distance on CLR-transformed data) matrix. We then performed a PCA on the Aitchison distance matrix to visually assess multivariate differences between samples ( Figure 12 A-C).
3.3.2 Betadispersion, PERMANOVA & Beta Diversity
Homogeneity of multivariate dispersion was assessed using the Aitchison distance matrix with the betadisper() function from the vegan package prior to conducting a PERMANOVA to determine if microbiome sample ASV composition differed significantly across developmental stage and leachate level.
PERMANOVA tests for differences in group centroids in multivariate space but assumes that within-group dispersion is comparable across groups. If dispersions differ significantly, PERMANOVA may detect differences driven by unequal variability rather than true shifts in community composition. By first evaluating whether average distances to group centroids differed across groups, we determined whether the assumption of homogeneous dispersion was met.
The permutation test for homogeneity of multivariate dispersions revealed no significant difference in dispersion across leachate (F3,59 = 0.14, p = 0.939, 999 permutations), stage (F2,60 = 1.0498, p = 0.36, 999 permutations), or the interaction between leachate and stage (F11,51 = 0.35, p = 0.971, 999 permutations), indicating groups have equal variance and meet the assumptions of PERMANOVA. However, we found that multivariate dispersion differed significantly across spawn nights (F2,60 = 6.29, p = 0.003, 999 permutations). Microbiome composition was clearly more varied on July 7th compared to July 6th and 8th, evident by the larger spread around the group centroid (Figure 11 A).
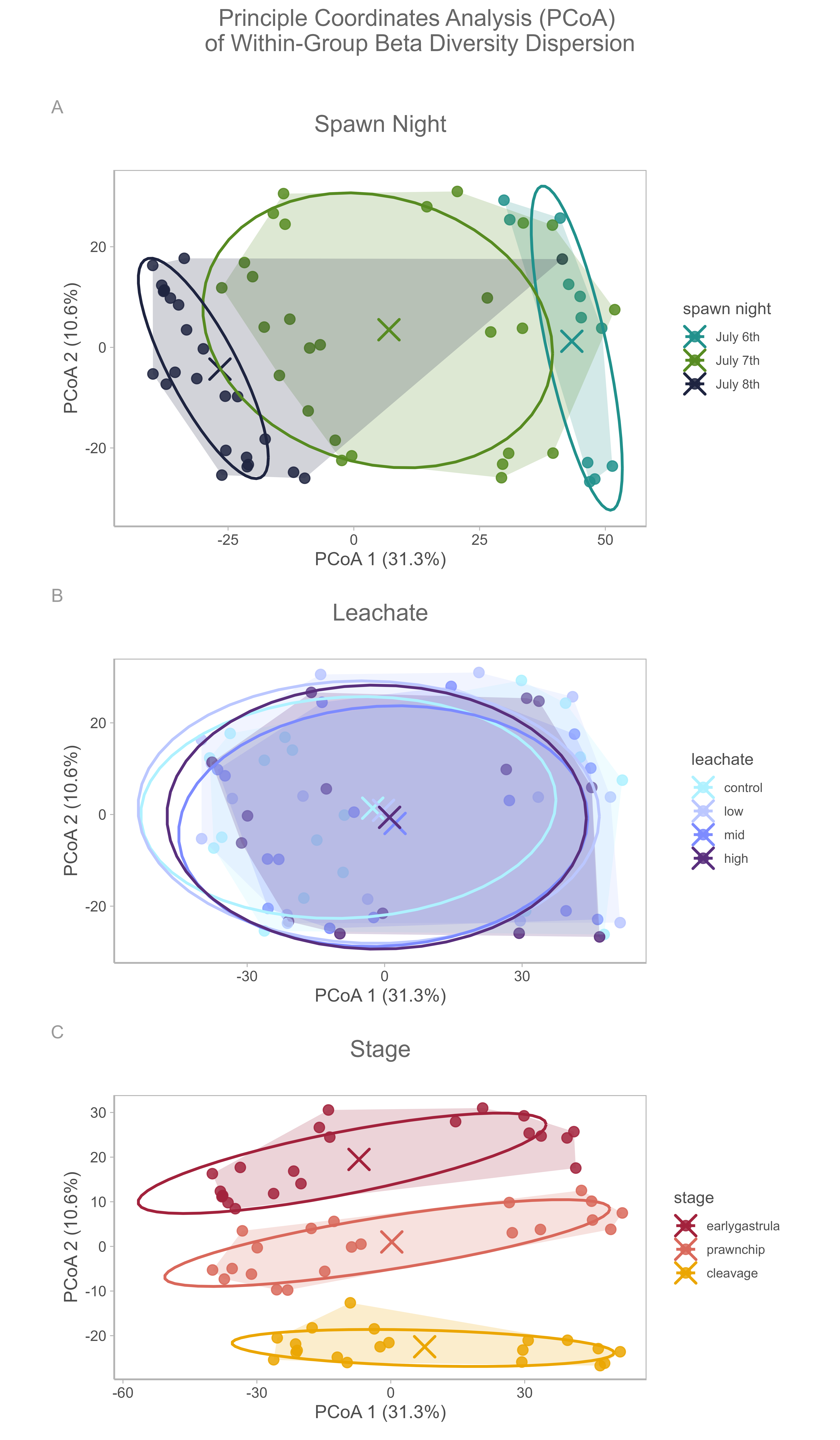
This finding prompted us to control for spawn night as a nuisance factor in our experiment by restricting permutations of samples among others within the same spawn night via strata, which accounts for the non-independence of samples collected on the same night when calculating the significance of our factors of interest (developmental stage and leachate level).
We conducted a community composition analysis using a PERMANOVA conducted on Aitchison distances with 999 permutations, stratified by spawn night ( Figure 12 A). It revealed a significant effect of developmental stage on community composition ( F2,51 = 3.84, R2 = 0.119, p = 0.001, 999 permutations), indicating that stage explained approximately 11.9% of the variation in multivariate structure (Figure 12 C). In contrast, leachate treatment had no significant effect ( F3,51 = 0.84, R2 = 0.039, p = 0.297, Figure 12 B), and there was no significant leachate × stage interaction ( F6,51 = 0.51, R2=0.048, p=1.000) .
3.3.3 Differentially abundant and prevalent bacterial taxa
To identify individual taxa associated with leachate exposure over developmental time, MaAsLin3 generalized linear mixed models were applied at the genus (L6) taxonomic level using the model formula “abundance|prevalence ~ leachate * hpf + reads + (1|spawn_night)”. This framework tested both abundance and prevalence responses while accounting for sequencing depth and variation among spawn nights.
Across 885 bacterial genera tested, eight genera showed significant associations with leachate exposure in the abundance model and eight genera showed significant associations in the prevalence model after filtering for models without fitting errors and joint false discovery rates of q < 0.1. Seven of these genera overlapped between the abundance and prevalence analyses, indicating that most responsive taxa exhibited coordinated shifts in both relative abundance and detection probability across treatments. Responsive taxa included members of the genera Tropicibacter, Sulfitobacter, Peredibacter, Pontibacterium, PS1 clade taxa, uncultured Neisseriaceae, and Marine Methylotrophic Group 3 ( Figure 13). One genus within the Longimicrobiaceae was uniquely significant in prevalence models ( Figure 14), while Leeuwenhoekiella was uniquely significant in abundance models ( Figure 13).
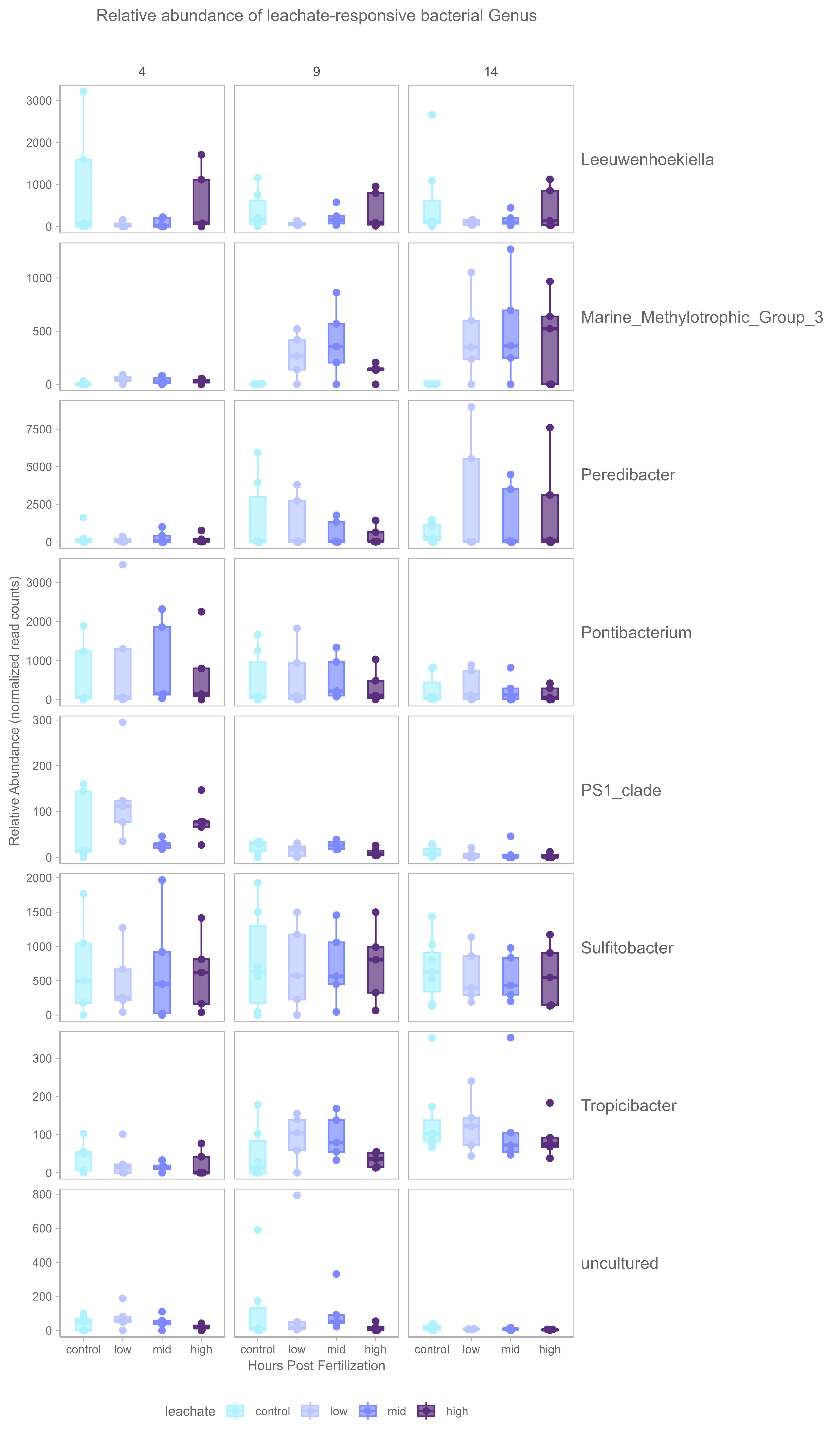
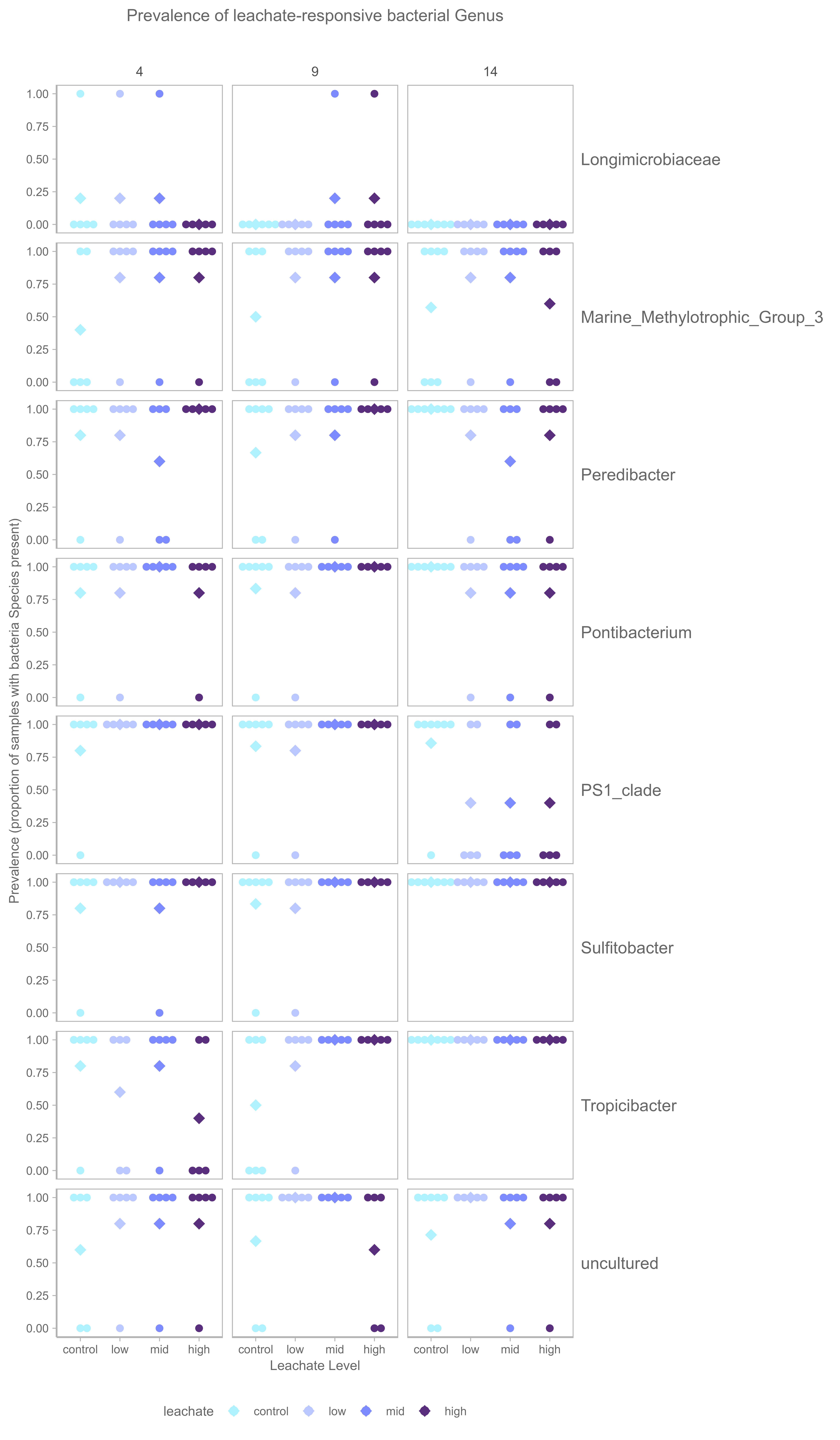
In contrast, an alternative model treating leachate level as a continuous linear predictor detected no significant leachate-associated taxa. The absence of significant linear responses, combined with significant associations in the categorical model, suggests that bacterial responses to PVC leachate were non-linear across exposure levels. Several taxa displayed the strongest shifts at low or intermediate leachate levels compared to the high leachate level, rather than a monotonic relationship across the level gradient, consistent with our hypothesis that we would observe low-dose responses.
Although only a small proportion of taxa responded significantly to leachate exposure (0.9% of genera tested), these results exceeded expectations under random chance given the applied false discovery threshold and stringent filtering criteria. Relative abundance and prevalence visualizations further indicated that responsive taxa varied across developmental stages, with some genera exhibiting stage-specific increases or decreases in abundance and detection probability under leachate exposure. Overall, these findings suggest that PVC leachate did not broadly restructure the embryonic microbiome, but instead produced targeted and non-linear shifts in a limited subset of bacterial taxa.
3.3.4 Predicted functional enrichment
Differential abundance and prevalence testing of PICRUSt2-predicted MetaCyc pathways identified multiple microbial functional pathways associated with PVC leachate exposure and stage-dependent responses. Functional shifts were distributed across pathways involved in methanogenesis, carbon fixation, aromatic compound degradation, chlorinated compound degradation, and precursor metabolite biosynthesis.
Several significant pathways were associated with methane metabolism and anaerobic carbon cycling, including the superpathway of C1 compound oxidation to CO2 (PWY-1882), methyl-coenzyme M oxidation to CO2 I (PWY-5209), and the reductive acetyl-CoA pathway in autotrophic methanogens (PWY-7784). These pathways are primarily associated with archaeal methanogenic and methylotrophic metabolism and suggest shifts in microbial functional potential related to anaerobic respiration and carbon processing in the Marine Methylotrophic Group 3 differentially abundant taxa.
Additional predicted pathways involved aromatic and chlorinated compound degradation, including cinnamate and hydroxycinnamate degradation (PWY-6690), 3-phenylpropanoate degradation (HCAMHPDEG-PWY; PWY0-1277), toluene degradation (TOLUENE-DEG-3-OH-PWY), pentachlorophenol degradation (PCPDEG-PWY), and dichlorobenzene degradation (14DICHLORBENZDEG-PWY; PWY-6084). Many of these pathways are associated with bacterial degradation of aromatic hydrocarbons and chlorinated compounds, which are commonly linked to anthropogenic pollution and industrial contaminants. The enrichment of pathways related to aromatic and chlorinated compound metabolism suggests that PVC leachate exposure may favor microbial taxa capable of metabolizing plastic-associated organic compounds or responding to chemically stressful environments.
Predicted pathways associated with lipid metabolism and signaling were also detected, including phosphatidate metabolism as a signaling molecule (PWY-7039). Because PICRUSt2 predicts functional potential from taxonomic composition rather than directly measuring gene expression or metabolism, these results represent inferred functional capacity rather than confirmed metabolic activity.
4 Discussion
Plastic pollution is an emerging threat to coral reefs, not only as a physical stressor but also as a source of chemical contaminants that may disrupt sensitive developmental processes. Here, we show that environmentally relevant levels of PVC leachate can trigger subtle shifts in developmental timing, gene expression, and specific microbial taxa in Montipora capitata within the first 14 hours of development, despite no detectable effects on survival or visible abnormalities. These findings reveal that plastic-derived chemicals can induce early, sublethal biological responses that are missed by traditional toxicity endpoints, highlighting previously under-explored mechanisms by which plastic pollution may already be impacting coral resilience.
4.1 H1: Coral development slightly accelerated at low and moderate PVC leachate exposures
The effects of leachate varied across both level and developmental time. The strongest treatment-dependent response was observed during the prawnchip stage, particularly under low and moderate PVC leachate exposure. These results indicate that PVC leachate did not broadly disrupt embryogenesis or induce developmental arrest, but instead caused subtle, stage-specific shifts in developmental pace. Developmental timing in marine larvae is highly plastic and responds to environmental conditions, such as temperature, with warmer conditions often accelerating embryonic development and influencing dispersal and recruitment success (O’Connor et al. 2007; Woolsey et al. 2013). Our findings suggest that plastic-derived chemicals may similarly influence developmental timing by interacting with endocrine-like signaling pathways involved in growth and differentiation (Tarrant (2007), Figure 8). The fact that responses were strongest at low and intermediate levels, rather than at the highest exposure level, is consistent with non-monotonic response patterns commonly reported for endocrine-disrupting compounds such as plasticizers and other PVC additives (Vandenberg et al. 2012).
One prior study found that weathered polypropylene microplastics minimally reduced fertilization success and increased embryo abnormalities in a non-dose dependent manner in the reef-building coral Acropora tenuis (Berry et al. 2019). Furthermore, exposure to high-density polyethylene (HDPE) leachate reduced larval survival and settlement in both M. capitata and Porites species, with higher levels (200mg/L) of high-density polyethylene (HDPE) leachate treatments paradoxically promoting settlement, potentially through attraction to chemical cues associated with degrading plastics (Wilkins and Richmond (2025)).
Our findings suggest that the prawnchip stage represents a critical developmental bottleneck in Montipora capitata, as embryos at this stage are more prone to fragmentation, likely due to the thin single-cell epithelial layer that characterizes this bizarre stage. We observed more torn embryos at 9 hpf relative to 4 or 14 hpf regardless of treatment (Figure 3 C), indicating that this stage may be particularly vulnerable to mechanical damage or developmental abnormalities. In some corals, embryo fragmentation has been proposed as a mechanism for increased reproductive output, provided that fragmented embryos remain viable and continue to develop as twins (Heyward and Negri 2012).
Our morpological findings are supported by our transcriptmoic data, which showed that leachate-responsive genes during early development were associated with cellular signaling and mitotic cell cycle processes (Figure 8, Figure 9), pathways that are central to regulating developmental progression.
Ramakrishnan and Wayne (2008) demonstrated that low-dose bisphenol A (BPA) exposure accelerated embryonic development, advanced hatching, and accelerated reproductive maturation in medaka (Oryzias latipes), with these effects mediated through thyroid hormone signaling pathways rather than classical estrogenic signaling. Similarly, Anselmo et al. (2011) reported that triphenyltin (TPT) and dibutyltin (DBT), two endocrine-disrupting chemicals commonly added to PVC products, accelerated metamorphosis in the sea urchin Psammechinus miliaris. Together, these studies suggest that plastic-associated endocrine-disrupting compounds can accelerate or influence developmental timing by interfering with signaling pathways that regulate growth and differentiation.
An alternative explanation is that low and moderate levels of PVC leachate exposure selectively favored embryos that were already developing more rapidly, resulting in an apparent shift in developmental progression rather than a true acceleration of development at the individual level. Distinguishing between true developmental acceleration and selective survival would require longitudinal tracking of individual embryos and larvae.
Notably, most previous studies examining plastic leachate effects on marine invertebrates have reported developmental disruption, arrest, or delay rather than accelerated development. For example, Paganos et al. (2023) documented severe embryotoxicity and morphological abnormalities in Strongylocentrotus purpuratus embryos exposed to PVC pellet leachate, with zinc identified as a primary driver of toxicity. Similarly, Jimenez-Guri et al. (2023) reported concentration-dependent disruption of axial formation, cell specification, and morphogenesis across multiple animal phyla, including cnidarians. However, the concentrations used in these studies were substantially higher than those tested in our study. It is possible that high levels of PVC leachate overwhelm subtle signaling-mediated effects on developmental timing responses and instead produce more pronounced toxic responses.
4.2 H2: PVC leachate impacted genes responsible for development, gene regulation, and cellular communication
Early coral embryogenesis is characterized by rapid cell division, gastrulation, and extensive use of maternally provisioned resources (Huffmyer et al. 2023; Chille et al. 2021). Our results showed that developmental stage explained substantially more transcriptomic variation than PVC leachate exposure. Rather than causing broad disruption of gene expression, PVC leachate induced comparatively subtle molecular responses embedded within the larger signal of gene expression across development. This pattern is consistent with developmental canalization, or the idea that early embryos may be partially buffered against environmental perturbations through maternal mRNA provisioning (Chille et al. 2021; Chille et al. 2022), allowing core developmental processes to remain stable despite exposure to complex contaminants.
Although overall gene expression remained largely unchanged, we identified 130 leachate-responsive genes associated with cellular signaling, transcriptional regulation, vesicle-mediated transport, lipid metabolism, and mitotic cell cycle activity (Figure 8), suggesting that PVC leachate primarily affected regulatory and cellular maintenance pathways rather than causing widespread developmental disruption. The stage-specific nature of these responses further indicates that sensitivity to leachate exposure changes across embryogenesis, likely reflecting differences in developmental state, energetic demand, or the timing of key cellular transitions. For example, cleavage-stage responses were associated with chromosomal and nuclear organization (Figure 9 A), whereas later gastrulation-stage responses were more strongly linked to cellular remodeling, signaling, and intracellular transport (Figure 9 C).
Among the leachate-responsive genes several were associated with G protein-coupled receptor (GPCR) signaling and related developmental signaling pathways. Similar patterns have also been observed in other cnidarians, where exposure of the sea anemone Exaiptasia diaphana to oxybenzone (a common sunscreen additive) and benzyl butyl phthalate (a plasticizer) altered the expression of candidate endocrine-disruption biomarker genes associated with GPCR-related signaling pathways (Morgan et al. 2022). This previous study highlighted genes involved in steroid hormone biosynthesis, cholesterol transport, immunity, phagocytosis, and Hedgehog signaling, including Niemann-Pick C type 2 (NPC2), Cathepsin L (CTSL), Smoothened (SMO), Desert Hedgehog (DHH), Zinc finger protein GLI2 (GLI2), and Vitellogenin (VTG). However, none of the leachate-responsive Montipora capitata transcripts mapped directly to the same genes identified by Morgan et al. (2022) in our study.
Building on these observations, we examined whether PVC leachate-responsive genes in Montipora capitata were enriched for GPCR-related signaling pathways. To do this we filtered DEGs for GO terms named GPCR signaling, signal transduction, receptor, transmembrane signaling, second messenger, hedgehog, adenylate cyclase, Wnt, and cell signaling. This analysis identified 74 GO terms associated with GPCR-related pathways, including regulation of GPCR signaling pathways, steroid hormone receptor signaling, nuclear receptor-mediated steroid hormone signaling, and canonical Wnt signaling. These GO terms mapped to 32 of the 130 leachate-responsive DEGs, including 6 genes associated with cleavage-stage expression patterns, 8 associated with prawnchip-stage patterns, and 16 associated with early gastrula-stage patterns.
GPCR signaling pathways are membrane-bound receptors that detect extracellular signals, such as peptides, hormones, or environmental cues and relay them intracellularly through G proteins and second messengers such as cyclic AMP and calcium signaling. These cascades regulate transcriptional programs involved in cell differentiation, morphogenesis, developmental timing, and stress responses (Grasso et al. 2008). The presence of GO terms associated with Wnt and Hedgehog signaling is particularly notable because these pathways are highly conserved regulators of embryonic axis formation, tissue patterning, and cell fate determination across metazoans (Pires-daSilva and Sommer 2003).
Although cnidarians lack centralized endocrine organs, they possess diverse peptide-mediated signaling systems that may function in endocrine-like regulation (Thiel et al. 2023; Twan et al. 2006). In cnidarians, neuropeptides and extracellular signaling molecules interact heavily with GPCR pathways to regulate development, environmental sensing, metabolism, and larval settlement (Grasso et al. 2001; Hambleton et al. 2018). Because GPCR pathways can amplify subtle extracellular signals into large downstream transcriptional responses through second messenger cascades (Civciristov et al. 2018), they may represent a plausible mechanism underlying the low-dose and non-monotonic transcriptional responses we observed in this study (Vandenberg et al. 2012).
Our findings suggest that plastic-derived chemicals may influence developmental regulation in Montipora capitata, potentially via endocrine-like cellular signaling pathways. Although cnidarians do not possess discrete endocrine organs comparable to those found in vertebrates, several components of steroid signaling pathways have been identified in corals and other anthozoans (Tarrant 2005; Tarrant 2007). In vertebrates, reproductive development and gametogenesis are coordinated through endocrine signaling networks in which hormones bind receptor proteins that regulate downstream gene transcription, and disruption of these pathways by exogenous chemicals can alter developmental timing and physiology (McLachlan 2001). Comparable endocrine-like signaling mechanisms may also occur in corals.
Our findings suggest that plastic-derived chemicals may influence developmental regulation in Montipora capitata, trhough endocrine-like cellular signaling pathways. Components of steroid signaling systems, including estradiol and estrone, have been detected in coral tissues of Montipora capitata and in seawater during spawning events (Tarrant et al. 1999; Slattery et al. 1999; Atkinson and Atkinson 1992; Twan et al. 2006). Corals are also capable of steroid metabolism (Tarrant et al. 2003), supporting the presence of hormonally responsive biochemical pathways that could potentially be disrupted by plastic-associated chemicals (Tarrant 2005).
The relatively targeted responses we observed in our study contrast with previous work showing broader developmental disruption following PVC leachate exposure. Paganos et al. (2023) reported extensive downregulation of developmental genes in purple sea urchin embryos exposed to PVC leachates, suggesting impairment of normal developmental progression. Differences in leachate composition, exposure concentration, exposure duration, or species-specific sensitivity may explain the more limited responses observed in our study. The absence of broad transcriptional disruption further suggests that the environmentally relevant concentrations used in our study remained below thresholds associated with acute developmental toxicity.
Our results show low and moderate PVC leachate levels elicited stronger transcriptional responses than the highest level, consistent with non-monotonic stress responses observed in other developmental systems. For example, environmentally relevant concentrations of Di(2-ethylhexyl) phthalate (DEHP), a common PVC plasticizer, produced biphasic responses in antioxidant activity and energy metabolism in Mytilus galloprovincialis, including U-shaped and inverted U-shaped dose-response relationships Xu et al. (2021). Together, these findings suggest that sublethal exposure to PVC leachate may not produce simple dose-dependent toxicity, but instead may alter developmental signaling in complex, non-monotonic ways. This pattern is consistent with the disruption of sensitive regulatory pathways, such as GPCR-mediated signaling.
4.3 H3: PVC leachate exposure produced targeted and non-linear shifts in a limited subset of bacterial taxa and may alter the functional potential of the embryonic coral microbiome
Corals depend on dynamic interactions with their associated microbial communities to support nutrient cycling, immune defense, and holobiont health (Glaze et al. 2022; Peixoto et al. 2017; Voolstra et al. 2024). In our study, embryonic microbiome composition was influences primarily by developmental progression rather than by PVC leachate exposure, consistent with previous work showing that bacterial communities in Montipora capitata shift across developmental stages (Huffmyer et al. 2025; Bernasconi et al. 2019).
Although PVC leachate did not broadly restructure the embryo-associated microbiome, it did alter the abundance and prevalence of specific bacterial taxa. In particular, the Methylophaga Marine Methylotrophic Group 3 increased consistently across all PVC leachate treatments.
Members of Methylophaga are obligate marine methylottrophs that play important roles in coral sulfur cycling by consuming dimethylsulfide (DMS), a breakdown product of dimethylsulfoniopropionate (DMSP), produced by corals and their symbiotic algae (Raina et al. 2013, 2010). DMSP and its derivatives contribute to antioxidant defense, stress signaling, and antimicrobial activity within the coral holobiont (Deschaseaux et al. 2014; Jackson et al. 2020; Gardner et al. 2017; Aguilar et al. 2017). The enrichment of Methylophaga under PVC leachate exposure may therefore reflect either utilization of plastic-derived methylated compounds or disruption of coral sulfur metabolism (Kim et al. 2007; Kröber and Schäfer 2019; Janvier and Grimont 1995; Neufeld et al. 2007).
In either case, increased methylotrophic activity could alter the embryo’s surrounding chemical environment (by accelerating DMS consumption) and potentially reduce antioxidant and stress-response capacity (Reisch et al. 2011; Fischer and Jones 2012).
Predicted functional profiling suggested enrichment of pathways associated with methanogenesis and the degradation of aromatic hydrocarbons and chlorinated compounds. These functional shifts are plausible because PVC leachates contain mixtures of complex dissolved organic carbon, including plasticizers, aromatic compounds, and chlorinated additives, which may favor specialized marine microbes capable of metabolizing them. Similar microbial metabolic restructuring has been reported in stressed coral microbiomes, including increases in sulfur and nitrogen cycling, virulence, and secondary metabolism (Vega Thurber et al. 2009). Enrichment of methanogenesis-associated pathways may reflect selection for microbes adapted to altered carbon cycling or capable of tolerating or metabolizing plastic-associated chemicals.
Likewise, enrichment of pathways related to aromatic hydrocarbon and chlorinated compound degradation is consistent with previous studies showing that coral-associated microbial communities can harbor bacteria capable of degrading hydrocarbons and other xenobiotic compounds (Al-Dahash and Mahmoud 2013; Villela et al. 2023).
Many reef-associated bacteria naturally metabolize halogenated compounds present in marine environments and may therefore be pre-adapted to tolerate or transform chlorinated chemicals released from PVC as leachate (Atashgahi et al. 2018).
These functional predictions should be interpreted with caution, as PICRUSt-based inference from 16S rRNA data strongly depends on the representation of reference genomes, which remains limited for many coral-associated marine microbes (Douglas et al. 2020). While these traits were inferred from amplicon sequence variant (ASV) phylogenetic placement rather than direct metagenomic or proteomic measurements, the results suggest that PVC leachate pollution may influence not only which microbes associate with developing coral embryos, but also the broader metabolic potential of the embryo-associated microbiome.
Our findings are also broadly consistent with previous marine microbiome studies showing that PVC leachates can restructure seawater microbial-communities (Focardi et al. 2022) and enrich genes associated with pathogenicity, motility, antibiotic resistence, and stress response (Focardi et al. 2022; Vlaanderen et al. 2023). Although these studies were not coral-specific, and did not identify Methylophaga, they support the broader conclusion that PVC leachates can alter the microbial environment surrounding corals and their developing embryos.
Together, these results suggest that environmentally relevant PVC exposure may exert subtle but ecologically meaningful selective pressures on microbial assembly during coral embryogenesis, even when overall community structure remains relatively stable.
5 Conclusion
At the surface, Montipora capitata coral embryos appear largely resilient to PVC leachate stress, yet we detected several subtle but important biological responses to PVC leachate pollution. Leachate exposure was associated with small, stage-specific shifts in developmental timing, suggesting that key embryonic transitions may be modestly accelerated by PVC leachate exposure. At the molecular level, 130 genes responded to leachate exposure, indicating targeted transcriptional effects nested within broader embryonic regulation. Similarly, the embryo-associated microbiome did not undergo large community-wide restructuring, yet specific bacterial taxa (and subsequently their predicted functional pathways) were impacted by PVC leachate exposure, particularly those linked to aromatic carbon degradation and DMSP cycling. Together, these findings suggest that PVC leachate may exert hidden sublethal effects that are not captured by traditional survival-based endpoints.
These findings reveal that exposure to plastic-derived chemicals can subtly alter development, gene regulation, and host-associated microbiomes during the most sensitive stages of coral life history. Such early, low-dose effects may scale up to influence larval performance, recruitment success, and ultimately coral population resilience. The patterns detected here are consistent with the expected effects of endocrine-disrupting chemicals operating through low-dose, endocrine-like signaling.
Future studies should investigate whether acute early-life exposure to PVC leachate produces longer-term consequences that persist beyond embryogenesis, including effects on epigenetic regulation, larval survival trajectories, metamorphosis, and settlement success. Because this study focused on acute exposure, additional work examining chronic or repeated exposure scenarios is also needed to better reflect environmentally realistic conditions. Plastic pollution rarely occurs in isolation in reef environments, and future experiments should therefore examine how PVC leachate interacts with additional stressors such as thermal stress from ocean warming and marine heatwaves. Furthermore, PVC leachate represents only one subset of microplastic polymers and associated additive chemicals, and substantially more work is needed to characterize the toxicity of other plastic types and chemical mixtures.
This study is relevant to the conservation and management of coral reef ecosystems, and can inform conservation, management, and policy decisions on plastics and their chemical additives as emerging contaminants of concern. Support for regulations that recognize and mitigate the impacts of low-dose endocrine-disrupting chemicals on vulnerable marine ecosystems is critical for protecting these vital life-giving resources (Vandenberg 2021).
6 Acknowledgements
This experiment was conducted on Oahu, Hawaiʻi. As guests, we recognize and give thanks for the land and water resources of the ʻaina and the traditional owners of the land, kanaka ʻoiwi, both past and present, as well as future generations, on which this experimental work was conducted in the Kaneʻohe Ahupuaʻa. Molecular lab work and computational analysis were conducted in Seattle, WA. We therefore wish to acknowledge the Coast Salish peoples of the land that touches the shared waters of all tribes and bands within the Suquamish, Tulalip, and Muckleshoot nations. We are grateful for the logistical support provided by the Donohue Lab, the Coral Resilience Lab and the Hawaiʻi Institute of Marine Biology. We acknowledge use of the computational resources of the Roberts Lab, and have immense gratitude for the guidance of Sam White and Ariana Huffmyer. We acknowledge coral colony collection by Callum Backstrom and Katherine Lasdin, as well as experimental assay setup and sample collection by Katherine Lasdin. We acknowledge Savanah Leidholdt, Jesse Zaneveld, and Amy Van Cise for helpful discussions on 16S bacterial analyses.
7 Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
9 Funding
This research was supported by the National Science Foundation CAREER Grant (award no. ), the University of Washington College of Environment Benjamin and Margaret Hall Student Research Support Fund , and the University of Washington School of Aquatic & Fishery Sciences Finishing Fellowship.
10 Data availability
Raw paired-end reads (FASTQ format) of RNA-Seq and 16S were deposited and are publicly available in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) under BioProject accession numbers PRJNA1177827 and PRJNA1244351.
11 AI
Declaration of generative AI and AI-assisted technologies in the manuscript preparation process
During the preparation of this work the author(s) used ChatGPT in order to summarize methods of analysis in R, and Elicit to search for peer reviewed papers. After using this tool/service, the author(s) reviewed and edited the content as needed and take(s) full responsibility for the content of the published article.
12 Supplementary
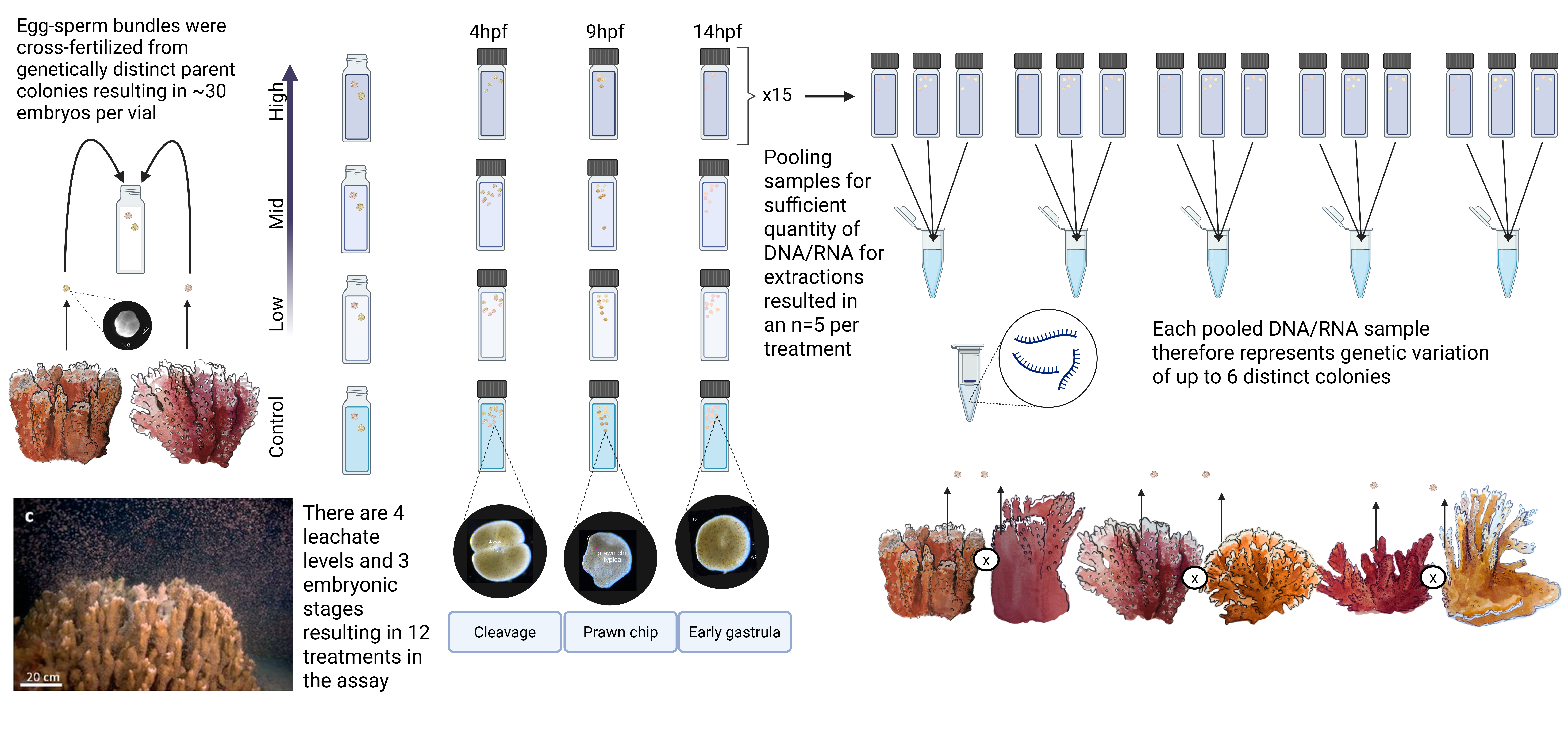

| Term | df | Sum Sq | Mean Sq | F value | Pr(>F) |
|---|---|---|---|---|---|
| PVC leachate treatment | 3 | 57.51852 | 19.17284 | 0.5228956 | 0.6675647 |
| Developmental stage (hpf) | 2 | 4696.35185 | 2348.17593 | 64.0411616 | 0.0000000 |
| Treatment × stage | 6 | 125.87037 | 20.97840 | 0.5721380 | 0.7516086 |
| Residuals | 96 | 3520.00000 | 36.66667 | NA | NA |
| Effect | Df | Deviance | p-value | Dev | p | Dev | p | Dev | p | Dev | p | Dev | p |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | 3 | 1.50 | 0.998 | 0.14 | 0.999 | 0.52 | 0.999 | 0.36 | 0.999 | 0.03 | 0.999 | 0.46 | 0.999 |
| hpf | 2 | 568.00 | 0.001** | 111.98 | 0.001** | 76.46 | 0.001** | 71.99 | 0.001** | 126.17 | 0.001** | 181.39 | 0.001** |
| Treatment × hpf | 6 | 40.30 | 0.009** | 16.53 | 0.108 | 2.76 | 0.144 | 9.42 | 0.108 | 11.59 | 0.108 | 0.00 | 0.791 |
| Effect | Df | Deviance | p-value | Dev | p | Dev | p | Dev | p |
|---|---|---|---|---|---|---|---|---|---|
| Treatment | 3 | 5.47 | 0.701 | 0.37 | 0.949 | 0.86 | 0.949 | 4.24 | 0.470 |
| hpf | 2 | 90.91 | 0.001** | 63.75 | 0.001** | 25.24 | 0.001** | 1.91 | 0.387 |
| Treatment × hpf | 6 | 12.12 | 0.886 | 4.69 | 0.870 | 6.14 | 0.854 | 1.29 | 0.982 |
References
Aguilar, Catalina, Jean-Baptiste Raina, Cherie A Motti, et al. 2017. “Transcriptomic Analysis of the Response of Acropora Millepora to Hypo-Osmotic Stress Provides Insights into DMSP Biosynthesis by Corals.” BMC Genomics 18 (August): 612. https://doi.org/10.1186/s12864-017-3959-0.
Al-Dahash, Lulwa M, and Huda M Mahmoud. 2013. “Harboring Oil-Degrading Bacteria: A Potential Mechanism of Adaptation and Survival in Corals Inhabiting Oil-Contaminated Reefs.” Marine Pollution Bulletin 72 (July): 364–74. https://doi.org/10.1016/j.marpolbul.2012.08.029.
Anselmo, Henrique M R, Lina Koerting, Sarah Devito, et al. 2011. “Early Life Developmental Effects of Marine Persistent Organic Pollutants on the Sea Urchin Psammechinus Miliaris.” Ecotoxicology and Environmental Safety 74 (November): 2182–92. https://doi.org/10.1016/j.ecoenv.2011.07.037.
Atashgahi, Siavash, Martin G Liebensteiner, Dick B Janssen, Hauke Smidt, Alfons J M Stams, and Detmer Sipkema. 2018. “Microbial Synthesis and Transformation of Inorganic and Organic Chlorine Compounds.” Frontiers in Microbiology 9 (December): 3079. https://doi.org/10.3389/fmicb.2018.03079.
Atkinson, S, and M J Atkinson. 1992. “Detection of Estradiol-17? During a Mass Coral Spawn.” Coral Reefs 11 (April): 33–35. https://doi.org/10.1007/bf00291932.
Bernasconi, Rachele, Michael Stat, Annette Koenders, Andrea Paparini, Michael Bunce, and Megan J Huggett. 2019. “Establishment of Coral-Bacteria Symbioses Reveal Changes in the Core Bacterial Community with Host Ontogeny.” Frontiers in Microbiology 10 (July): 1529. https://doi.org/10.3389/fmicb.2019.01529.
Berry, Kathryn L E, Hannah E Epstein, Phoebe J Lewis, Nora M Hall, and Andrew P Negri. 2019. “Microplastic Contamination Has Limited Effects on Coral Fertilisation and Larvae.” Diversity 11 (November): 228. https://doi.org/10.3390/d11120228.
Bolyen, Evan, Jai Ram Rideout, Matthew R Dillon, et al. 2019. “Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2.” Nature Biotechnology 37 (August): 852–57. https://doi.org/10.1038/s41587-019-0209-9.
Capolupo, Marco, Ayesha Rafiq, Irene Coralli, et al. 2023. “Bioplastic Leachates Characterization and Impacts on Early Larval Stages and Adult Mussel Cellular, Biochemical and Physiological Responses.” Environmental Pollution (Barking, Essex: 1987) 319 (February): 120951. https://doi.org/10.1016/j.envpol.2022.120951.
Chen, Shifu. 2023. “Ultrafast One-Pass FASTQ Data Preprocessing, Quality Control, and Deduplication Using Fastp.” iMeta 2: e107. https://doi.org/10.1002/imt2.107.
Chille, E E, E L Strand, F Scucchia, et al. 2022. “Energetics, but Not Development, Is Impacted in Coral Embryos Exposed to Ocean Acidification.” The Journal of Experimental Biology 225 (October): jeb243187. https://doi.org/10.1242/jeb.243187.
Chille, Erin, Emma Strand, Mayaan Neder, et al. 2021. “Developmental Series of Gene Expression Clarifies Maternal mRNA Provisioning and Maternal-to-Zygotic Transition in a Reef-Building Coral.” BMC Genomics 22 (November): 815. https://doi.org/10.1186/s12864-021-08114-y.
Civciristov, Srgjan, Andrew M Ellisdon, Ryan Suderman, et al. 2018. “Preassembled GPCR Signaling Complexes Mediate Distinct Cellular Responses to Ultralow Ligand Concentrations.” Science Signaling 11 (October): eaan1188. https://doi.org/10.1126/scisignal.aan1188.
Claar, Danielle C, Samuel Starko, Kristina L Tietjen, et al. 2020. “Dynamic Symbioses Reveal Pathways to Coral Survival Through Prolonged Heatwaves.” Nature Communications 11 (December): 6097. https://doi.org/10.1038/s41467-020-19169-y.
Danecek, Petr, James K Bonfield, Jennifer Liddle, et al. 2021. “Twelve Years of SAMtools and BCFtools.” GigaScience 10 (February). https://doi.org/10.1093/gigascience/giab008.
Deschaseaux, E S M, G B Jones, M A Deseo, et al. 2014. “Effects of Environmental Factors on Dimethylated Sulfur Compounds and Their Potential Role in the Antioxidant System of the Coral Holobiont.” Limnology and Oceanography 59 (May): 758–68. https://doi.org/10.4319/lo.2014.59.3.0758.
Douglas, Gavin M, Vincent J Maffei, Jesse R Zaneveld, et al. 2020. “PICRUSt2 for Prediction of Metagenome Functions.” Nature Biotechnology 38 (June): 685–88. https://doi.org/10.1038/s41587-020-0548-6.
Eddy, Tyler D, William W L Cheung, and John F Bruno. 2018. “Historical Baselines of Coral Cover on Tropical Reefs as Estimated by Expert Opinion.” PeerJ 6 (January): e4308. https://doi.org/10.7717/peerj.4308.
Eddy, Tyler D, Vicky W Y Lam, Gabriel Reygondeau, et al. 2021. “Global Decline in Capacity of Coral Reefs to Provide Ecosystem Services.” One Earth (Cambridge, Mass.) 4 (September): 1278–85. https://doi.org/10.1016/j.oneear.2021.08.016.
Ewels, Philip, Måns Magnusson, Sverker Lundin, and Max Käller. 2016. “MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report.” Bioinformatics (Oxford, England) 32 (October): 3047–48. https://doi.org/10.1093/bioinformatics/btw354.
Fischer, Esther, and Graham Jones. 2012. “Atmospheric Dimethysulphide Production from Corals in the Great Barrier Reef and Links to Solar Radiation, Climate and Coral Bleaching.” Biogeochemistry 110 (September): 31–46. https://doi.org/10.1007/s10533-012-9719-y.
Focardi, Amaranta, Lisa R Moore, Jean-Baptiste Raina, Justin R Seymour, Ian T Paulsen, and Sasha G Tetu. 2022. “Plastic Leachates Impair Picophytoplankton and Dramatically Reshape the Marine Microbiome.” Microbiome 10 (October): 179. https://doi.org/10.1186/s40168-022-01369-x.
Gambardella, Chiara, Roberta Miroglio, Marina Prieto Amador, et al. 2024. “High Concentrations of Phthalates Affect the Early Development of the Sea Urchin Paracentrotus Lividus.” Ecotoxicology and Environmental Safety 279 (July): 116473. https://doi.org/10.1016/j.ecoenv.2024.116473.
Gardner, Stephanie G, Jean-Baptiste Raina, Matthew R Nitschke, et al. 2017. “A Multi-Trait Systems Approach Reveals a Response Cascade to Bleaching in Corals.” BMC Biology 15 (December): 117. https://doi.org/10.1186/s12915-017-0459-2.
Glaze, Thomas D, Dirk V Erler, and Henri M P Siljanen. 2022. “Microbially Facilitated Nitrogen Cycling in Tropical Corals.” The ISME Journal 16 (January): 68–77. https://doi.org/10.1038/s41396-021-01038-1.
Gloor, Gregory B, Jean M Macklaim, Vera Pawlowsky-Glahn, and Juan J Egozcue. 2017. “Microbiome Datasets Are Compositional: And This Is Not Optional.” Frontiers in Microbiology 8 (November): 2224. https://doi.org/10.3389/fmicb.2017.02224.
Grasso, L C, D C Hayward, J W Trueman, K M Hardie, P A Janssens, and E E Ball. 2001. “The Evolution of Nuclear Receptors: Evidence from the Coral Acropora.” Molecular Phylogenetics and Evolution 21 (October): 93–102. https://doi.org/10.1006/mpev.2001.0994.
Grasso, Lauretta C, John Maindonald, Stephen Rudd, et al. 2008. “Microarray Analysis Identifies Candidate Genes for Key Roles in Coral Development.” BMC Genomics 9 (November): 540. https://doi.org/10.1186/1471-2164-9-540.
Gunaalan, Kuddithamby, Elena Fabbri, and Marco Capolupo. 2020. “The Hidden Threat of Plastic Leachates: A Critical Review on Their Impacts on Aquatic Organisms.” Water Research 184 (October): 116170. https://doi.org/10.1016/j.watres.2020.116170.
Hahladakis, John N, Costas A Velis, Roland Weber, Eleni Iacovidou, and Phil Purnell. 2018. “An Overview of Chemical Additives Present in Plastics: Migration, Release, Fate and Environmental Impact During Their Use, Disposal and Recycling.” Journal of Hazardous Materials 344 (February): 179–99. https://doi.org/10.1016/j.jhazmat.2017.10.014.
Hambleton, Elizabeth, Victor A S Jones, Ira Maegele, David Vaskoff, Timo Sachsenheimer, and Annika Guse. 2018. “Enhanced Stability of Non-Canonical NPC2 in the Symbiosome Supports Coral-Algal Symbiosis.” bioRxiv, August 25, 399766. https://doi.org/10.1101/399766.
Hermabessiere, Ludovic, Alexandre Dehaut, Ika Paul-Pont, et al. 2022. “Occurrence and Effects of Plastic Additives on Marine Environments and Organisms: A Review.” Chemosphere 182 (February): 781–93. https://doi.org/10.1016/j.chemosphere.2017.05.096.
Heyward, A J, and A P Negri. 2012. “Turbulence, Cleavage, and the Naked Embryo: A Case for Coral Clones.” Science (New York, N.Y.) 335 (March): 1064. https://doi.org/10.1126/science.1216055.
Huffmyer, Ariana S, Kevin H Wong, Danielle M Becker, Emma Strand, Tali Mass, and Hollie M Putnam. 2023. “Multi-Omic Analysis of Coral Development Elucidates the Metabolic Transitions from Parental Provisioning to Endosymbiotic Nutritional Exchange.” bioRxiv, ahead of print, March 23. https://doi.org/10.1101/2023.03.20.533475.
Huffmyer, Ariana S, Kevin H Wong, Danielle M Becker, Emma Strand, Tali Mass, and Hollie M Putnam. 2025. “Shifts and Critical Periods in Coral Metabolism Reveal Energetic Vulnerability During Development.” Current Biology 35 (June): 2858–2871.e6. https://doi.org/10.1016/j.cub.2025.05.013.
Isa, Valerio, Francesco Saliu, Alessandro Becchi, et al. 2025. “Impacts of Microplastics on Reef-Building Corals: Disentangling the Contribution of the Chain Scission Products Released by Weathering.” The Science of the Total Environment 975 (May): 179239. https://doi.org/10.1016/j.scitotenv.2025.179239.
Isa, Valerio, Davide Seveso, Luca Diamante, et al. 2024. “Physical and Cellular Impact of Environmentally Relevant Microplastic Exposure on Thermally Challenged Pocillopora Damicornis (Cnidaria, Scleractinia).” The Science of the Total Environment 918 (February): 170651. https://doi.org/10.1016/j.scitotenv.2024.170651.
Jackson, Rebecca L, Albert J Gabric, Roger Cropp, and Matthew T Woodhouse. 2020. “Dimethylsulfide (DMS), Marine Biogenic Aerosols and the Ecophysiology of Coral Reefs.” Biogeosciences 17 (April): 2181–204. https://doi.org/10.5194/bg-17-2181-2020.
Jambeck, Jenna R, Roland Geyer, Chris Wilcox, et al. 2015. “Marine Pollution. Plastic Waste Inputs from Land into the Ocean.” Science (New York, N.Y.) 347 (February): 768–71. https://doi.org/10.1126/science.1260352.
Janvier, M, and P A Grimont. 1995. “The Genus Methylophaga, a New Line of Descent Within Phylogenetic Branch Gamma of Proteobacteria.” Research in Microbiology 146 (September): 543–50. https://doi.org/10.1016/0923-2508(96)80560-2.
Jimenez-Guri, Eva, Carola Murano, Periklis Paganos, and Maria Ina Arnone. 2023. “PVC Pellet Leachates Affect Adult Immune System and Embryonic Development but Not Reproductive Capacity in the Sea Urchin Paracentrotus Lividus.” Marine Pollution Bulletin 196 (November): 115604. https://doi.org/10.1016/j.marpolbul.2023.115604.
Kim, Daehwan, Joseph M Paggi, Chanhee Park, Christopher Bennett, and Steven L Salzberg. 2019. “Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype.” Nature Biotechnology 37 (August): 907–15. https://doi.org/10.1038/s41587-019-0201-4.
Kim, Hee Gon, Nina V Doronina, Yuri A Trotsenko, and Si Wouk Kim. 2007. “Methylophaga Aminisulfidivorans Sp. Nov., a Restricted Facultatively Methylotrophic Marine Bacterium.” International Journal of Systematic and Evolutionary Microbiology 57 (September): 2096–101. https://doi.org/10.1099/ijs.0.65139-0.
Kröber, Eileen, and Hendrik Schäfer. 2019. “Identification of Proteins and Genes Expressed by Methylophaga Thiooxydans During Growth on Dimethylsulfide and Their Presence in Other Members of the Genus.” Frontiers in Microbiology 10 (May): 1132. https://doi.org/10.3389/fmicb.2019.01132.
Lamb, Joleah B, Bette L Willis, Evan A Fiorenza, et al. 2018. “Plastic Waste Associated with Disease on Coral Reefs.” Science (New York, N.Y.) 359 (January): 460–62. https://doi.org/10.1126/science.aar3320.
Langfelder, Peter, and Steve Horvath. 2008. “WGCNA: An R Package for Weighted Correlation Network Analysis.” BMC Bioinformatics 9 (December): 559. https://doi.org/10.1186/1471-2105-9-559.
Law, R J, T W Fileman, and P Matthiessen. 1991. “Phthalate Esters and Other Industrial Organic Chemicals in the North and Irish Seas.” Water Science and Technology: A Journal of the International Association on Water Pollution Research 24 (November): 127–34. https://doi.org/10.2166/wst.1991.0283.
Love, Michael I, Wolfgang Huber, and Simon Anders. 2014. “Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2.” Genome Biology 15 (December): 550. https://doi.org/10.1186/s13059-014-0550-8.
Lynch, Jennifer M, Katrina Knauer, and Katherine R Shaw. 2022. Plastic Additives in the Ocean. 1st ed. Edited by Anthony L Andrady. Wiley. https://doi.org/10.1002/9781119768432.ch2.
MacLeod, Matthew, Hans Peter H Arp, Mine B Tekman, and Annika Jahnke. 2021. “The Global Threat from Plastic Pollution.” Science (New York, N.Y.) 373 (July): 61–65. https://doi.org/10.1126/science.abg5433.
Mallick, Himel, Ali Rahnavard, Lauren J McIver, et al. 2021. “Multivariable Association Discovery in Population-Scale Meta-Omics Studies.” PLoS Computational Biology 17 (November): e1009442. https://doi.org/10.1371/journal.pcbi.1009442.
Maqbool, Faheem, Sara Mostafalou, Haji Bahadar, and Mohammad Abdollahi. 2016. “Review of Endocrine Disorders Associated with Environmental Toxicants and Possible Involved Mechanisms.” Life Sciences 145 (January): 265–73. https://doi.org/10.1016/j.lfs.2015.10.022.
Mathieu, C, and J Bednarek. 2022. Survey of Phthalates in Washington State Waterbodies, 2021. Research Report Nos. 22-03-027. Washington State Department of Ecology, Olympia. https://apps.ecology.wa.gov/publications/documents/2203027.pdf.
McLachlan, J A. 2001. “Environmental Signaling: What Embryos and Evolution Teach Us about Endocrine Disrupting Chemicals.” Endocrine Reviews 22 (June): 319–41. https://doi.org/10.1210/edrv.22.3.0432.
Morgan, Michael B, James Ross, Joseph Ellwanger, et al. 2022. “Sea Anemones Responding to Sex Hormones, Oxybenzone, and Benzyl Butyl Phthalate: Transcriptional Profiling and in Silico Modelling Provide Clues to Decipher Endocrine Disruption in Cnidarians.” Frontiers in Genetics 12. https://www.frontiersin.org/articles/10.3389/fgene.2021.793306.
Net, Sopheak, Anne Delmont, Richard Sempéré, Andrea Paluselli, and Baghdad Ouddane. 2015. “Reliable Quantification of Phthalates in Environmental Matrices (Air, Water, Sludge, Sediment and Soil): A Review.” The Science of the Total Environment 515-516 (May): 162–80. https://doi.org/10.1016/j.scitotenv.2015.02.013.
Neufeld, Josh D, Hendrik Schäfer, Michael J Cox, Rich Boden, Ian R McDonald, and J Colin Murrell. 2007. “Stable-Isotope Probing Implicates Methylophaga Spp and Novel Gammaproteobacteria in Marine Methanol and Methylamine Metabolism.” The ISME Journal 1 (October): 480–91. https://doi.org/10.1038/ismej.2007.65.
O’Connor, Mary I, John F Bruno, Steven D Gaines, et al. 2007. “Temperature Control of Larval Dispersal and the Implications for Marine Ecology, Evolution, and Conservation.” Proceedings of the National Academy of Sciences of the United States of America 104 (January): 1266–71. https://doi.org/10.1073/pnas.0603422104.
Okubo, Nami, Takuma Mezaki, Yoko Nozawa, et al. 2013. “Comparative Embryology of Eleven Species of Stony Corals (Scleractinia).” PloS One 8 (December): e84115. https://doi.org/10.1371/journal.pone.0084115.
Oppen, Madeleine J H van, Ruth D Gates, Linda L Blackall, et al. 2017. “Shifting Paradigms in Restoration of the World’s Coral Reefs.” Global Change Biology 23 (September): 3437–48. https://doi.org/10.1111/gcb.13647.
Padilla-Gamiño, J L, T M Weatherby, R G Waller, and R D Gates. 2011. “Formation and Structural Organization of the Egg–Sperm Bundle of the Scleractinian Coral Montipora Capitata.” Coral Reefs 30 (June): 371–80. https://doi.org/10.1007/s00338-010-0700-8.
Padilla-Gamiño, and Gates. 2012. “Spawning Dynamics in the Hawaiian Reef-Building Coral Montipora Capitata.” Marine Ecology Progress Series 449 (March): 145–60. https://doi.org/10.3354/meps09530.
Paganos, Periklis, Clemens Vinzenz Ullmann, Daniela Gaglio, et al. 2023. “Plastic Leachate-Induced Toxicity During Sea Urchin Embryonic Development: Insights into the Molecular Pathways Affected by PVC.” The Science of the Total Environment 864 (March): 160901. https://doi.org/10.1016/j.scitotenv.2022.160901.
Paluselli, Andrea, Vincent Fauvelle, Natascha Schmidt, François Galgani, Sopheak Net, and Richard Sempéré. 2018. “Distribution of Phthalates in Marseille Bay (NW Mediterranean Sea).” The Science of the Total Environment 621 (April): 578–87. https://doi.org/10.1016/j.scitotenv.2017.11.306.
Peixoto, Raquel S, Phillipe M Rosado, Deborah Catharine de Assis Leite, Alexandre S Rosado, and David G Bourne. 2017. “Beneficial Microorganisms for Corals (BMC): Proposed Mechanisms for Coral Health and Resilience.” Frontiers in Microbiology 8. https://www.frontiersin.org/article/10.3389/fmicb.2017.00341.
Pertea, Mihaela, Geo M Pertea, Corina M Antonescu, Tsung-Cheng Chang, Joshua T Mendell, and Steven L Salzberg. 2015. “StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads.” Nature Biotechnology 33 (March): 290–95. https://doi.org/10.1038/nbt.3122.
Pinheiro, Marlene, Irene Martins, Joana Raimundo, Miguel Caetano, Teresa Neuparth, and Miguel M Santos. 2023. “Stressors of Emerging Concern in Deep-Sea Environments: Microplastics, Pharmaceuticals, Personal Care Products and Deep-Sea Mining.” The Science of the Total Environment 876 (June): 162557. https://doi.org/10.1016/j.scitotenv.2023.162557.
Pires-daSilva, André, and Ralf J Sommer. 2003. “The Evolution of Signalling Pathways in Animal Development.” Nature Reviews. Genetics 4 (January): 39–49. https://doi.org/10.1038/nrg977.
Raina, Jean-Baptiste, Elizabeth A Dinsdale, Bette L Willis, and David G Bourne. 2010. “Do the Organic Sulfur Compounds DMSP and DMS Drive Coral Microbial Associations?” Trends in Microbiology 18 (March): 101–8. https://doi.org/10.1016/j.tim.2009.12.002.
Raina, Jean-Baptiste, Dianne M Tapiolas, Sylvain Forêt, et al. 2013. “DMSP Biosynthesis by an Animal and Its Role in Coral Thermal Stress Response.” Nature 502 (October): 677–80. https://doi.org/10.1038/nature12677.
Ramakrishnan, Siddharth, and Nancy L Wayne. 2008. “Impact of Bisphenol-a on Early Embryonic Development and Reproductive Maturation.” Reproductive Toxicology (Elmsford, N.Y.) 25 (February): 177–83. https://doi.org/10.1016/j.reprotox.2007.11.002.
Reaka-Kudla, Marjorie L. 1997. “The Global Biodiversity of Coral Reefs: A Comparison with Rainforests.” In Biodiversity II: Understanding and Protecting Our Biological Resources, edited by Marjorie L. Reaka-Kudla, Don E. Wilson, and Edward O. Wilson. National Academies Press. https://www.researchgate.net/publication/239063261.
Reisch, Chris R, Mary Ann Moran, and William B Whitman. 2011. “Bacterial Catabolism of Dimethylsulfoniopropionate (DMSP).” Frontiers in Microbiology 2 (August): 172. https://doi.org/10.3389/fmicb.2011.00172.
Rendell-Bhatti, Flora, Periklis Paganos, Anna Pouch, et al. 2021. “Developmental Toxicity of Plastic Leachates on the Sea Urchin Paracentrotus Lividus.” Environmental Pollution (Barking, Essex: 1987) 269 (January): 115744. https://doi.org/10.1016/j.envpol.2020.115744.
Richmond, Robert H. 1997. “Reproduction and Recruitment in Corals: Critical Links in the Persistence of Reefs.” In Life and Death of Coral Reefs. Springer US. https://doi.org/10.1007/978-1-4615-5995-5_8.
Richmond, Robert H, Kaho H Tisthammer, and Narrissa P Spies. 2018. “The Effects of Anthropogenic Stressors on Reproduction and Recruitment of Corals and Reef Organisms.” Frontiers in Marine Science 5 (July): 379626. https://doi.org/10.3389/fmars.2018.00226.
Rochman, Chelsea M, Cole Brookson, Jacqueline Bikker, et al. 2019. “Rethinking Microplastics as a Diverse Contaminant Suite.” Environmental Toxicology and Chemistry 38 (April): 703–11. https://doi.org/10.1002/etc.4371.
Saliu, Francesco, Simone Montano, Marina Lasagni, and Paolo Galli. 2020. “Biocompatible Solid-Phase Microextraction Coupled to Liquid Chromatography Triple Quadrupole Mass Spectrometry Analysis for the Determination of Phthalates in Marine Invertebrate.” Journal of Chromatography A 1618 (May): 460852. https://doi.org/10.1016/j.chroma.2020.460852.
Slattery, M, G A Hines, J Starmer, and V J Paul. 1999. “Chemical Signals in Gametogenesis, Spawning, and Larval Settlement and Defense of the Soft Coral Sinularia Polydactyla.” Coral Reefs 18 (April): 75–84. https://doi.org/10.1007/s003380050158.
Souter, David, Serge Planes, Jérémy Wicquart, Murray Logan, David Obura, and Francis Staub. 2020. Status of Coral Reefs of the World: 2020. Research report. Global Coral Reef Monitoring Network & International Coral Reef Initiative. https://gcrmn.net/wp-content/uploads/2021/10/Executive-Summary-with-Forewords.pdf.
Stephens, Timothy G, Junmo Lee, Yujin Jeong, et al. 2022. “High-Quality Genome Assembles from Key Hawaiian Coral Species.” GigaScience 11 (November): giac098. https://doi.org/10.1093/gigascience/giac098.
Stubbins, Aron, Kara Lavender Law, Samuel E Muñoz, Thomas S Bianchi, and Lixin Zhu. 2021. “Plastics in the Earth System.” Science (New York, N.Y.) 373 (July): 51–55. https://doi.org/10.1126/science.abb0354.
Tarrant, A M. 2005. “Endocrine-Like Signaling in Cnidarians: Current Understanding and Implications for Ecophysiology.” Integrative and Comparative Biology 45 (January): 201–14. https://doi.org/10.1093/icb/45.1.201.
Tarrant, A M, M J Atkinson, and S Atkinson. 2004. “Effects of Steroidal Estrogens on Coral Growth and Reproduction.” Marine Ecology Progress Series 269: 121–29. https://doi.org/10.3354/meps269121.
Tarrant, A M, S Atkinson, and M J Atkinson. 1999. “Estrone and Estradiol-17 Beta Concentration in Tissue of the Scleractinian Coral, Montipora Verrucosa.” Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology 122 (January): 85–92. https://doi.org/10.1016/s1095-6433(98)10155-1.
Tarrant, Ann M. 2007. “Hormonal Signaling in Cnidarians: Do We Understand the Pathways Well Enough to Know Whether They Are Being Disrupted?” Ecotoxicology (London, England) 16 (February): 5–13. https://doi.org/10.1007/s10646-006-0121-1.
Tarrant, Ann M, C H Blomquist, P H Lima, M J Atkinson, and S Atkinson. 2003. “Metabolism of Estrogens and Androgens by Scleractinian Corals.” Comparative Biochemistry and Physiology. Part B, Biochemistry & Molecular Biology 136 (November): 473–85. https://doi.org/10.1016/s1096-4959(03)00253-7.
Tetu, Sasha G, Indrani Sarker, Verena Schrameyer, et al. 2019. “Plastic Leachates Impair Growth and Oxygen Production in Prochlorococcus, the Ocean’s Most Abundant Photosynthetic Bacteria.” Communications Biology 2 (May): 184. https://doi.org/10.1038/s42003-019-0410-x.
Thiel, Daniel, Luis Alfonso Yañez-Guerra, Amanda Kieswetter, et al. 2023. “Large-Scale Deorphanization of Nematostella Vectensis Neuropeptide GPCRs Supports the Independent Expansion of Bilaterian and Cnidarian Peptidergic Systems.” bioRxiv, 2023.07.03.547448. https://doi.org/10.1101/2023.07.03.547448.
Twan, Wen-Hung, Jiang-Shiou Hwang, Yan-Horn Lee, Hua-Fang Wu, Ying-Hsiu Tung, and Ching-Fong Chang. 2006. “Hormones and Reproduction in Scleractinian Corals.” Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology 144 (July): 247–53. https://doi.org/10.1016/j.cbpa.2006.01.011.
Vandenberg, Laura N. 2021. “Endocrine Disrupting Chemicals: Strategies to Protect Present and Future Generations.” Expert Review of Endocrinology & Metabolism 16 (May): 135–46. https://doi.org/10.1080/17446651.2021.1917991.
Vandenberg, Laura N, Theo Colborn, Tyrone B Hayes, et al. 2012. “Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses.” Endocrine Reviews 33 (June): 378–455. https://doi.org/10.1210/er.2011-1050.
Vega Thurber, Rebecca, Dana Willner-Hall, Beltran Rodriguez-Mueller, et al. 2009. “Metagenomic Analysis of Stressed Coral Holobionts.” Environmental Microbiology 11 (August): 2148–63. https://doi.org/10.1111/j.1462-2920.2009.01935.x.
Vered, Gal, and Noa Shenkar. 2022. “Limited Effects of Environmentally-Relevant Concentrations in Seawater of Dibutyl Phthalate, Dimethyl Phthalate, Bisphenol a, and 4-Nonylphenol on the Reproductive Products of Coral-Reef Organisms.” Environmental Pollution (Barking, Essex: 1987) 314 (December): 120285. https://doi.org/10.1016/j.envpol.2022.120285.
Villela, Helena, Flúvio Modolon, Júnia Schultz, et al. 2023. “Genome Analysis of a Coral-Associated Bacterial Consortium Highlights Complementary Hydrocarbon Degradation Ability and Other Beneficial Mechanisms for the Host.” Scientific Reports 13 (July): 12273. https://doi.org/10.1038/s41598-023-38512-z.
Vlaanderen, Eric, Timothy Ghaly, Lisa Moore, Amaranta Focardi, Ian Paulsen, and Sasha Tetu. 2023. “Plastic Leachate Exposure Drives Antibiotic Resistance and Virulence in Marine Bacterial Communities.” bioRxiv, February 14, 2023.02.13.528379. https://doi.org/10.1101/2023.02.13.528379.
Voolstra, Christian R, Jean-Baptiste Raina, Melanie Dörr, et al. 2024. “The Coral Microbiome in Sickness, in Health and in a Changing World.” Nature Reviews. Microbiology 22 (August): 460–75. https://doi.org/10.1038/s41579-024-01015-3.
Wang, Yi, Ulrike Naumann, Stephen T Wright, and David I Warton. 2012. “Mvabund– an R Package for Model‐based Analysis of Multivariate Abundance Data: The Mvabund R Package.” Methods in Ecology and Evolution 3 (June): 471–74. https://doi.org/10.1111/j.2041-210x.2012.00190.x.
Weis, Judith S, and Karl H Palmquist. 2021. “Reality Check: Experimental Studies on Microplastics Lack Realism.” Applied Sciences (Basel, Switzerland) 11 (September): 8529. https://doi.org/10.3390/app11188529.
Wickham, Hadley, Mara Averick, Jennifer Bryan, et al. 2019. “Welcome to the Tidyverse.” Journal of Open Source Software 4 (November): 1686. https://doi.org/10.21105/joss.01686.
Wilkins, Keiko W, and Robert H Richmond. 2025. “Unseen Threats: Negative Effects of Microplastic Leachate on Coral Planulae Settlement.” Frontiers in Marine Science 12 (July): 1596594. https://doi.org/10.3389/fmars.2025.1596594.
Wilkins, Keiko W, Joanne Y Yew, Meredith Seeley, and Robert H Richmond. 2024. “Microplastic Leachate Negatively Affects Fertilization in the Coral Montipora Capitata.” Integrative and Comparative Biology 64 (October): 1131–40. https://doi.org/10.1093/icb/icae143.
Woolsey, E S, M Byrne, and A H Baird. 2013. “The Effects of Temperature on Embryonic Development and Larval Survival in Two Scleractinian Corals.” Marine Ecology Progress Series 493 (November): 179–84. https://doi.org/10.3354/meps10499.
Xu, Hai’e, Wanshuang Cao, Hongliang Sun, et al. 2021. “Dose-Dependent Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) in Mussel Mytilus Galloprovincialis.” Frontiers in Marine Science 8 (May): 658361. https://doi.org/10.3389/fmars.2021.658361.
Zhang, Ying, Yaqi Jiao, Zixu Li, Yue Tao, and Yang Yang. 2021. “Hazards of Phthalates (PAEs) Exposure: A Review of Aquatic Animal Toxicology Studies.” The Science of the Total Environment 771 (June): 145418. https://doi.org/10.1016/j.scitotenv.2021.145418.
Citation
BibTeX citation:
@online{tanja2026,
author = {Tanja, Sarah and Roberts, Steven and Saliu, Francesco and
Salipante, Stephan and L. Padilla-Gamiño, Jacqueline},
title = {Hidden {Impacts} of {Plastic} {Pollution} on {Coral}
{Embryos}},
date = {2026-06-03},
langid = {en},
abstract = {Ocean plastic pollution releases chemical compounds as
materials degrade. Polyvinyl Chloride (PVC) in particular contains
high levels of endocrine-disrupting additives that are ubiquitously
released into coastal environments as leachate. Despite this, the
impacts of PVC leachate on marine life remain poorly understood. We
investigated the ecotoxicological responses of three levels of PVC
leachate (0.01 mg/L, 0.1 mg/L, 1 mg/L) during the first 14 hours
post fertilization (hpf) of embryonic development in the coral
*Montipora capitata*. PVC leachate did not affect embryo survival or
the rate of morphological abnormality; however, we did detect subtle
changes in developmental timing. Furthermore, RNAseq analysis
identified 130 differentially expressed genes (DEGs) responsive to
PVC leachate treatments. Gene expression responses to leachate
exposure were strongest at low (0.01 mg/L) and moderate (0.1 mg/L)
levels, and during the earliest developmental stage (cleavage, 4
hpf), indicating a non-monotonic response pattern and suggesting
sublethal effects of low-dose PVC leachate exposure in developing
*Montipora capitata* embryos. Additionally, analysis of 16S rRNA
gene sequences (V4 region) revealed 9 bacterial taxa and 17
predicted microbial functional pathways responsive to PVC leachate,
with differences in abundance and/or prevalence compared to
controls. Our findings indicate that current environmental levels of
PVC pollution may already subtly influence gene expression and early
microbiome assembly in coral embryos as early as 4 hours
post-fertilization, providing new insights into the sublethal,
mechanistic impacts of plastic-derived chemicals.}
}
For attribution, please cite this work as:
Tanja, Sarah, Steven Roberts, Francesco Saliu, Stephan Salipante, and
Jacqueline L. Padilla-Gamiño. 2026. “Hidden Impacts of Plastic
Pollution on Coral Embryos.” UW Masters Thesis, June 3.